| 【亲耕田】植物学报特邀综述 | 您所在的位置:网站首页 › 绥梗18是叶上穗还是叶下穗 › 【亲耕田】植物学报特邀综述 |
【亲耕田】植物学报特邀综述
|
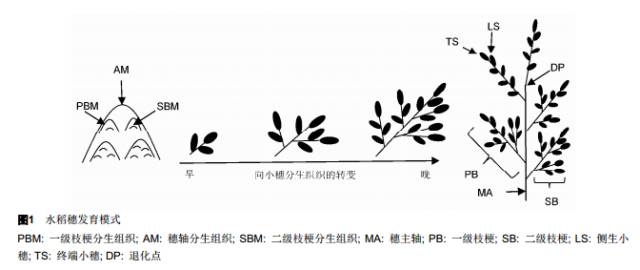
1 水稻穗部发育过程 水稻花序的主轴上着生多级枝梗, 枝梗长度由基部向顶端逐渐缩短, 使得整个花序呈圆锥状, 因此水稻花序亦称圆锥花序。水稻幼穗形成及发育期是决定小穗数目及最终产量的重要时期, 众多植物学家和农学家提出了多种对幼穗发育时期进行划分的方法。国内的经典划分方法是丁颖等(1959)提出的8个时期划分方式, 而国际上普遍接受的是9个时期的划分方式(Ikeda et al., 2004; Itoh et al.,2005)。
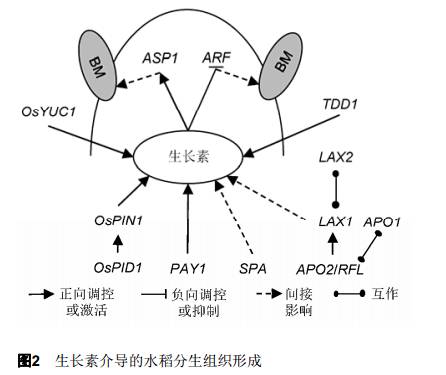
2 水稻穗型的遗传调控 水稻的穗分枝数直接决定着穗粒数, 而穗粒数又是与水稻产量相关的关键因素。因此, 研究调控水稻穗分枝形成机制不仅能解决植物发育相关的基本问题, 还对作物产量的提高有着重要作用。与模式植物拟南芥(Arabidopsis thaliana)的花序分枝不同, 水稻穗分枝的产生更为错综复杂。从一个单独的顶端分生组织到成穗, 众多基因参与调控, 进而形成诸如直立穗、弯曲穗、密穗、稀穗、长穗和短穗等千姿百态的穗型。从当前已有研究来看, 参与水稻幼穗发育的基因可分为4类, 分别调控枝梗分生组织的形成、枝梗分生组织的大小、小穗分生组织的转变时间以及枝梗的伸长。 2.1 生长素介导的分生组织形成 在禾本科的生殖分生组织中, 生长素(auxin)通过控制细胞极性和细胞伸长调控腋生分生组织的形成和外向生长(Cheng and Zhao, 2007; Zhao, 2010) (图2)。研究表明, 拟南芥中生长素合成酶YUC (YUCCA)、生长素转运蛋白PIN1 (PIN-FORMED 1)和PID(PINOID)以及生长素响应因子(ARF)在水稻中的同源物, 既是保守的又有不同之处(Benjamins and Scheres, 2008; Zhao, 2010), 其区别主要表现在编码此类蛋白的基因时空表达不同。
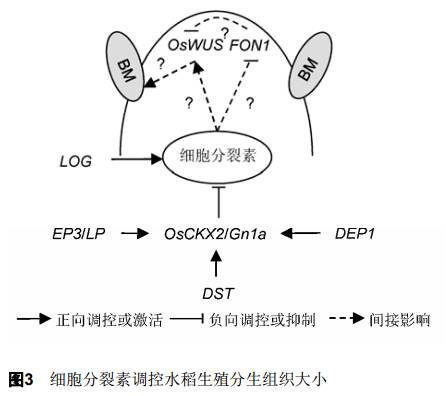
2.2 细胞分裂素调控生殖分生组织大小 细胞分裂素(cytokinins)能够促进细胞分裂, 在调控生殖分生组织大小和活性中起着基础与保守的作用,影响着单子叶植物和双子叶植物的分枝(Kyozuka,2007; Zhao, 2008) (图3)。由于细胞分裂素的代谢和信号能够通过调节生殖分生组织大小提高籽粒产量,因此将控制细胞分裂素水平的策略应用到育种方案中具有重要意义。
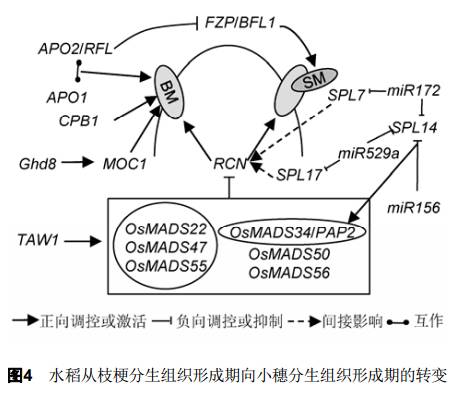
2.3 枝梗分生组织形成期向小穗分生组织形成期的转变 水稻花序从枝梗分生组织形成转入小穗形成是水稻穗型塑造中的重要事件, 这取决于分生组织的身份特性。通过对模式植物拟南芥的研究已发现了多个与分生组织身份确定相关的基因, 如对花分生组织身份进行协同调控的基因LFY (LEAFY)、 AP1 (APETALA1)和CAL (CAULIFLOWER), 这些基因使得侧生分生组织拥有花的身份特性进而向花分化(Mandel et al.,1992; Weigel et al., 1992; Kempin et al., 1995;Ferrándiz et al., 2000)。近年来在水稻中也发现了大量与穗部分生组织身份特性相关的基因, 此类基因变异往往导致小穗分生组织形成提前或延迟发生(图4)。
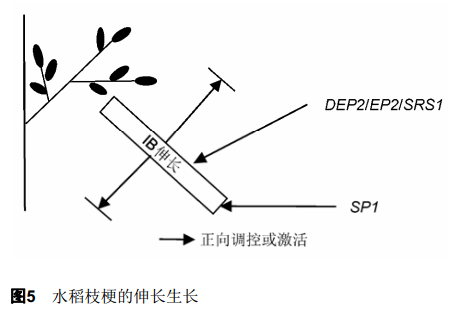
2.4 枝梗伸长的遗传调控 针对水稻枝梗伸长的研究目前还非常有限, 相关基因有DEP2 (Dense and Erect Panicle 2)和SP1 (Short Panicle 1) (图5)。此类基因突变后水稻幼穗各阶段分生组织形成正常, 但最终枝梗变短导致穗型变小。
2.5 水稻穗蘖分枝的协同调控 水稻分蘖和穗分枝分别是营养生长期和生殖生长期形成的侧生器官。某些情况下, 二者受到共同的调控,产生同等改变。如MOC1 (MONOCULM 1)和LAX1都参与分蘖和穗分枝, 其中任一基因突变都会导致穗蘖分枝数减少(Komatsu et al., 2003a; Li et al., 2003)。但多数情况下, 分蘖数和穗分枝数的改变并不总是保持一致, 如OsCKX2、 SP1和DEP1突变能特异地改变水稻穗分枝数, 分蘖数却未表现出相应变化(Ashikari et al., 2005; Huang et al., 2009; Li et al., 2009)。甚至在某些情况下还会出现相反的情况, 如NIL-ipa1植株表现为分蘖减少而穗分枝数增加(Li et al., 2010)。从一些与激素相关基因, 如Gnla (OsCKX2) (Ashikari et al., 2005)和D27 (Lin et al., 2009)的研究推测,穗蘖分枝的时空调控可能由不同的基因来完成。以上研究表明, 水稻在营养生长时的分蘖和生殖生长阶段的穗发育可能受不同的机制调控。
3 问题与展望 水稻穗型是与产量直接相关的性状, 对穗型的研究结果具有重要的应用前景, 已经成为众多分子生物学家和育种学家重点研究的方向之一。过去数十年来, 对控制水稻花序表型的相关因子及调控路径的研究已取得巨大进展, 如能特异地作用于水稻花序发育的生长素信号和细胞分裂素信号。一些开花时间调控因子也能影响水稻穗发育, 如Ghd8 (Grain number, Plant height, and Heading date 8)能促进MOC1的表达(Yan et al., 2011), 而MOC1所编码的GRAS家族核蛋白控制着水稻分蘖和穗分枝, 决定着分蘖数和花序一级枝梗与二级枝梗数(Oikawa and Kyozuka,2009)。生殖生长期, 水稻与双子叶植物拟南芥有着共同的调控机制, 包括CLV-WUS信号、生长素和细胞分裂素路径和MADS-box基因。尽管相关基因与路径在水稻中已被发现, 但其作用的具体模式仍有诸多未知之处, 未来应对此进行深入研究, 以期为水稻高产育种提供充分的理论依据。激素是调控植物生长发育的重要因素, 而植物株型调控机制多是通过内源激素调节分枝原基的产生与伸长。当前, 对植物株型塑造(即分枝)相关激素研究较多的有生长素、细胞分裂素和独脚金内酯(strigolactones)。独脚金内酯作为新的植物激素近年来才被发现, 该激素参与抑制植物分枝的伸长生长,同时与生长素和细胞分裂素一起调控植物的分枝数量。但目前已知的关于独脚金内酯的突变体或基因大多与分蘖相关, 如D3(Ishikawa et al., 2005)、 D14(Arite et al., 2009)和D27 (Lin et al., 2009)等, 尚未发现其在穗分枝发育中的作用。作为控制分枝发育的重要激素, 独脚金内酯是否参与调控水稻穗型及其相关机理还有待研究。影响穗发育的D11等位基因CPB1参与油菜素内酯(brassinolide)的合成, 然而油菜素内酯对水稻花序发育的调控作用尚未见相关报道。以上研究表明, 植物激素参与的水稻花序发育调控仍有众多未知。
当前, 突变体鉴定仍是研究水稻基因功能最为直接有效的方法。多年以来, 水稻相关生物学家和育种专家已发现众多有价值的变异材料, 如叶形突变体(鞠培娜等, 2010)、株高突变体(杨德卫等, 2011)及穗型突变体(刘丹等, 2016)等。而在已获得的众多穗型突变体中, 只有少数几个是功能获得的有利变异, 如大穗突变体lp (Li et al., 2011)。如何深入挖掘此类有利等位变异并应用到育种当中必然是将来的重大课题。当前获得变异资源的途径主要有2个, 一是自然变异, 二是人工诱变。针对自然变异, 要扩大相关种质资源的探索范围, 在一些地方品种中或许存在着更优质、更丰富的等位变异。而随着现代生物技术的不断发展, 利用新技术(如近年来兴起的CRISPR/Cas9)创制新的变异类型, 可大大丰富穗型的种质资源。
|






 返回搜狐,查看更多
返回搜狐,查看更多【本文地址】