| 在开发孢子脯氨酸合成所需的花粉发育和生育 | 您所在的位置:网站首页 › 致命的孢子怎么合成出来 › 在开发孢子脯氨酸合成所需的花粉发育和生育 |
在开发孢子脯氨酸合成所需的花粉发育和生育
|
启动子GydF4y2BaP5CS1.GydF4y2Ba和GydF4y2BaP5CS2.GydF4y2Ba在花粉发育后期的小孢子和花粉粒中有活性吗GydF4y2Ba
作为建立花粉发育和育性所需脯氨酸合成的第一步,我们分析了在花粉发育和育性控制下GUS酶活性的模式GydF4y2BaP5CS1.GydF4y2Ba和GydF4y2BaP5CS2.GydF4y2Ba在不同发展阶段的拟南芥中的启动子区域(图。GydF4y2Ba1GydF4y2Ba,附加文件GydF4y2Ba1GydF4y2Ba:图S1)。作为启动子序列,我们采用2932 bp的上游GydF4y2BaP5CS1.GydF4y2Ba和2097年的BP上游GydF4y2BaP5CS2.GydF4y2Ba起始密码子,根据Ábrahám等人(2003)[GydF4y2Ba29GydF4y2Ba].我们选择并分析了18个独立的变压器GydF4y2BaP.GydF4y2BaP5CS1.GydF4y2Ba:格斯GydF4y2Ba构建和21GydF4y2BaP.GydF4y2BaP5CS2.GydF4y2Ba:格斯。GydF4y2Ba从每个结构中,我们选择3个具有代表性染色模式的纯合子系进行更详细的分析。这两个GydF4y2BaP.GydF4y2BaP5CS1.GydF4y2Ba:格斯GydF4y2Ba和GydF4y2BaP.GydF4y2BaP5CS2.GydF4y2Ba:格斯GydF4y2Ba在花药中诱导特异性GUS表达,在主干中并在花梗中延伸到较小的延伸,但不是在鲜花或芽的其他部分中(图。GydF4y2Ba1GydF4y2Baa和h)。组织学横截面的分析显示GydF4y2BaP.GydF4y2BaP5CS1.GydF4y2Ba:格斯GydF4y2Ba和GydF4y2BaP.GydF4y2BaP5CS2.GydF4y2Ba:格斯GydF4y2Ba在花粉发育的第10 - 13阶段,即第一次有丝分裂之前到成熟花粉释放之前,小孢子和成熟花粉粒的发育特别活跃(图2)。GydF4y2Ba1GydF4y2Bab, c, d, e, f和g, i, j, k, l, m和n)。GydF4y2Ba1GydF4y2Bab和c, i和j),或在花药的任何孢子体组织中(图。GydF4y2Ba1GydF4y2Ba这些结果表明,拟南芥从第10期开始,花药中脯氨酸的合成几乎只发生在雄性配子体组织,即小孢子和花粉粒中,而在绒毡层、中间层和内膜等周围的孢子体组织中很少或不表达。GydF4y2Ba 图。1GydF4y2Ba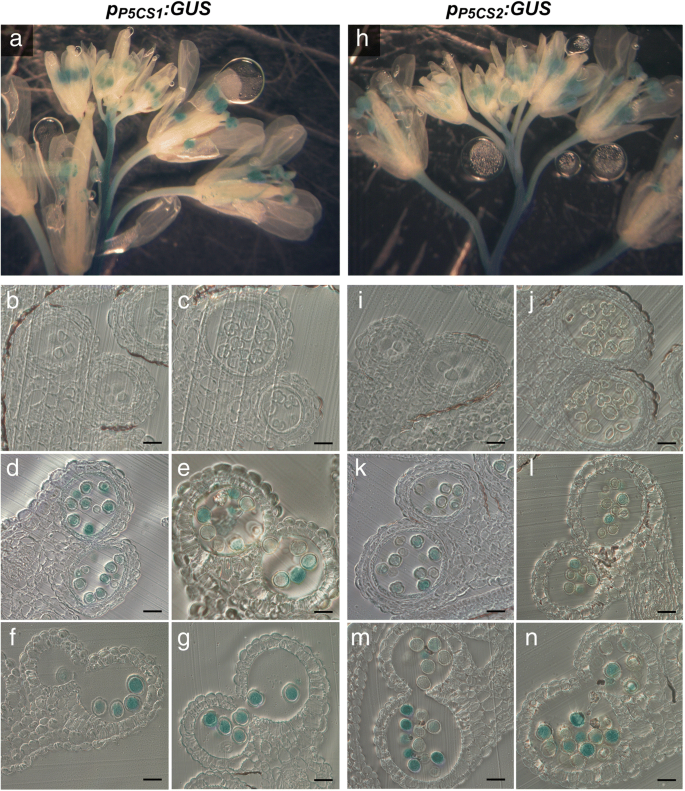
花药中Gus活性的组织化学定位GydF4y2BaP.GydF4y2BaP5CS1.GydF4y2Ba:格斯GydF4y2Ba和GydF4y2BaP.GydF4y2BaP5CS2.GydF4y2Ba:格斯GydF4y2Ba转基因拟南芥。花序的GydF4y2BaP.GydF4y2BaP5CS1.GydF4y2Ba:格斯GydF4y2Ba(GydF4y2BagGydF4y2Ba) 和GydF4y2BaP.GydF4y2BaP5CS2.GydF4y2Ba:格斯GydF4y2Ba(GydF4y2Bah nGydF4y2Ba)转基因植株用X-Gluc溶液浸润,染色过夜,固定后清除,进行显微分析。GydF4y2Ba一个hGydF4y2Ba花蕾和花在不同发育阶段的全山花序几乎只在花药中显示GUS活性。GydF4y2Bab-gGydF4y2Ba和GydF4y2Ba在GydF4y2Ba不同发育阶段的花药横切面:GydF4y2Bab,我GydF4y2Ba第8阶段与人蛋前微孔母细胞;GydF4y2Bac jGydF4y2Ba第9阶段小孢子四分体;GydF4y2Bad、kGydF4y2Ba第十阶段,两者都有强烈的活动GydF4y2BaP.GydF4y2BaP5CS1.GydF4y2Ba:格斯GydF4y2Ba和GydF4y2BaP.GydF4y2BaP5CS2.GydF4y2Ba:格斯GydF4y2Ba四处分离后;GydF4y2Bae、lGydF4y2Ba第11期具发育完全的外壁和退化的绒毡层;GydF4y2BaF,MGydF4y2Ba12阶段12带隔膜变性和GydF4y2BaG,N.GydF4y2Ba第13阶段与成熟花粉在花药裂开。在任何孢子素的花药组织中,几乎没有GUS染色。所有刻度条=20μmGydF4y2Ba 全尺寸图像GydF4y2Ba生物信息学启动子分析,包括基因本体论(GO)富集分析,表明在启动子中GydF4y2BaP5CS2.GydF4y2Ba并且,在较小程度上,GydF4y2BaP5CS1.GydF4y2Ba推定GydF4y2BaCIS.GydF4y2Ba-调控元件富集于与花粉发育和育性、花粉管生长、花药发育和双受精形成合子和胚乳相关的转录因子结合位点(附加文件)GydF4y2Ba2GydF4y2Ba:图S2)。此外,两者的发起人GydF4y2BaP5CS1.GydF4y2Ba和GydF4y2BaP5CS2.GydF4y2Ba包含WRKY34和WRKY2,参与花粉发育和功能[充分表征的转录因子识别位点GydF4y2Ba30.GydF4y2Ba那GydF4y2Ba31GydF4y2Ba].GydF4y2Ba Camv35s.GydF4y2Ba启动子驱动的孢子体表达GydF4y2BaP5CS2.GydF4y2Ba不拯救异常GydF4y2Bap5cs1 / p5cs2GydF4y2Ba花粉GydF4y2Ba在尝试补充异常花粉发育GydF4y2Bap5csGydF4y2Ba过度表达的癫痫GydF4y2BaP5CS2.GydF4y2Ba,我们引入了转基因的GydF4y2BaP5CS2.GydF4y2Ba在…的控制之下GydF4y2BaCamv35s.GydF4y2Ba据据报道,促进剂在拟南芥的大多数组织和器官中促进基因表达,尽管不适于发育梭菌和花粉颗粒[GydF4y2Ba32GydF4y2Ba].以确定其组织特异性GydF4y2BaCamv35s.GydF4y2Ba推动者,我们分析了携带A的植物的花药中的GUS活动GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba组织化学染色构建。在整个花序GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba在雌蕊,长丝和萼片中的衍射GUS活性以及不同发育阶段的花芽(图。GydF4y2Ba2GydF4y2Ba一个,附加文件GydF4y2Ba3.GydF4y2Ba:图S3A,B)。花树的组织学横截面GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba在维管束显示强GUS活性(图GydF4y2Ba2GydF4y2BaC)光和漫射活性GydF4y2BaCamv35s.GydF4y2Ba在减数分裂后的第8期,在花药和小孢子的所有孢子体组织中都有启动子(图。GydF4y2Ba2GydF4y2BaB,C和D)。在第9阶段,在无核梭菌中和阶段11和12的花粉晶粒中,没有检测到GUS活性(图。GydF4y2Ba2GydF4y2BaE和f),确认Wilkinson等人的结果。(1997)[GydF4y2Ba32GydF4y2Ba].GydF4y2Ba 图2GydF4y2Ba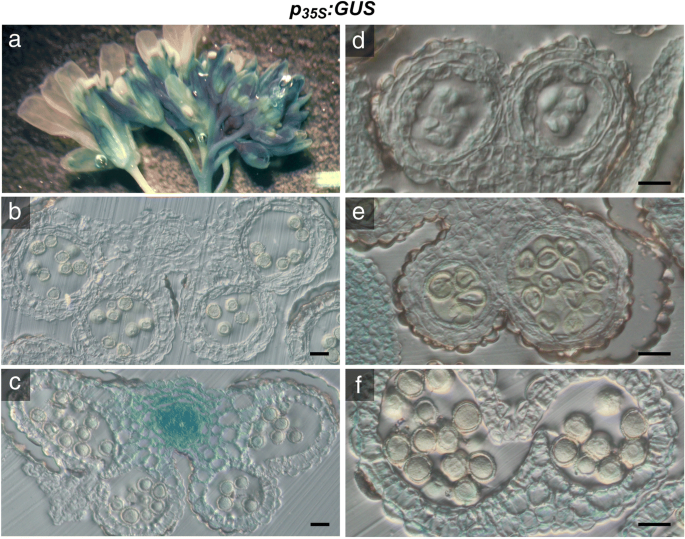
花药中Gus活性的组织化学定位GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba转基因拟南芥植物。花序的GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba将转基因植株用X-Gluc溶液浸润,染色过夜,固定并清除,进行显微分析。GydF4y2Ba一种GydF4y2Ba整个花序GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba在不同发育阶段的细丝,萼片,雌蕊和花芽中染色GUS染色。GydF4y2Bab-fGydF4y2Ba组织切片GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba花药矩:GydF4y2BaB.GydF4y2Ba未染色的花药在11/12阶段,由于光散射而显示出微弱的蓝色彩虹色。GydF4y2BaCGydF4y2Ba强大的GUS染色在第11/12阶段的血管束。在花粉囊内完全成熟的花粉中没有检测到GUS活性。酒吧在(GydF4y2BaB.GydF4y2Ba) 和 (GydF4y2BaCGydF4y2Ba= 50 μm。GydF4y2BaD.GydF4y2Ba第8期花药在高倍镜下显示弱,弥漫GUS染色在所有孢子体组织。GydF4y2BaE.GydF4y2Ba第9期花药在单核小孢子中无GUS染色。GydF4y2BaFGydF4y2Ba第12期花药在退化的花药壁中GUS活性较弱,而在成熟花粉中则没有。在一些花粉粒中可见的淡绿色可能表明35S启动子的残留活性,但也可能来自X-Gluc染色的可溶性中间体的扩散或花粉粒固有的低gus样活性[GydF4y2Ba57GydF4y2Ba那GydF4y2Ba58GydF4y2Ba那GydF4y2Ba59GydF4y2Ba].柱= 20 μm in (GydF4y2Bad-fGydF4y2Ba)GydF4y2Ba 全尺寸图像GydF4y2Ba随后,一个GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba结构被引入GydF4y2Bap5csGydF4y2Ba倍半突变体来评估组织表达的影响GydF4y2BaP5CS2.GydF4y2Ba在亲本孢子体组织中的发育GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变体花粉。但是,尽管存在存在GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba结果表明,在花序中检测到低水平的游离脯氨酸(图。GydF4y2Ba3.GydF4y2Ba一个,附加文件GydF4y2Ba4.GydF4y2Ba:表S1)GydF4y2Ba.GydF4y2Ba高水平GydF4y2BaP5CS2.GydF4y2Ba在植物营养发育的早期至开花期均有脯氨酸的表达和积累,而在开花前期则有脯氨酸的表达和积累GydF4y2BaP5CS2.GydF4y2Ba游离脯氨酸的表达量和水平下降到与原脯氨酸相似的水平GydF4y2Bap5csGydF4y2Ba昏迷线,表明GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba转基因被沉默(数据未显示)。在先前的研究中,我们观察到,轻度盐应激废除内源性的共抑制GydF4y2BaP5CSGydF4y2Ba转基因诱导的基因GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS1GydF4y2Ba构建[GydF4y2Ba9.GydF4y2Ba].因此,盐处理(0.1M NaCl)GydF4y2Bap5csGydF4y2Ba举行的戏曲仿真GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba构建恢复脯氨酸合成至类似于非应激野生型植物的水平(图。GydF4y2Ba3.GydF4y2Ba一个,附加文件GydF4y2Ba4.GydF4y2Ba:表S1)。然而,GUS酶活性的变化规律GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:格斯GydF4y2Ba0.1 M NaCl处理对花药的结构没有影响(数据未显示)。GydF4y2Ba 图3.GydF4y2Ba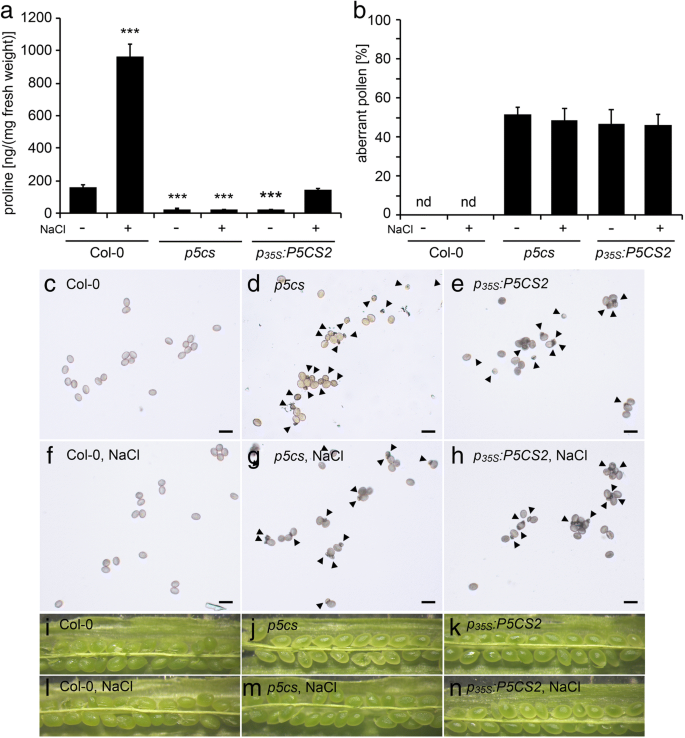
水稻花粉发育及育性分析GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba线条。GydF4y2Ba一种GydF4y2Ba野生型(Col-0)的花序中的脯氨酸积聚,GydF4y2Bap5csGydF4y2BaSesquimutants(GydF4y2Bap5csGydF4y2Ba) 和GydF4y2Bap5csGydF4y2Ba纯合的倍半突变体GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba构建(GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba).根据材料和方法,植株要么正常浇水,要么暴露在100mm NaCl下。条形图代表来自不同植物的至少三个样品的平均±SE。***表示与Col-0野生型有显著差异(GydF4y2BaP.GydF4y2Ba B.GydF4y2Ba从植物处理的植物中的花粉粒的百分比以与(GydF4y2Ba一种GydF4y2Ba).条形图代表52至311的±SE分析来自至少三种独立植物的花粉粒。nd:未检测到。GydF4y2Ba碳氢键GydF4y2Ba野生型花粉粒的亮视野显微照片(GydF4y2Bac、fGydF4y2Ba),GydF4y2Bap5csGydF4y2Ba(GydF4y2Bad、gGydF4y2Ba), 和GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba(GydF4y2Bae、hGydF4y2Ba)从正常浇水的植物(GydF4y2Bac, d, eGydF4y2Ba)或经NaCl处理的植物(GydF4y2BaF,G,HGydF4y2Ba).箭头表示小的、异常的花粉粒。柱= 50 μm。GydF4y2Ba在GydF4y2Ba开,过早的角果Stereomicroscopic图像。没有异常种子,指示的杀伤力GydF4y2Bap5cs2-1GydF4y2Ba纯合的胚胎(与图1相比较。GydF4y2Ba6.GydF4y2Ba),但在正常情况下均不可见(GydF4y2Ba我知道GydF4y2Ba)NaCl治疗后(GydF4y2BaL-M.GydF4y2Ba)GydF4y2Ba 全尺寸图像GydF4y2Ba无论是在正常条件下还是在轻度盐胁迫处理下,两者之间的差异均无统计学意义GydF4y2Bap5csGydF4y2Ba倍半植物有或没有GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba转基因,既不处于畸形花粉的频率(图。GydF4y2Ba3.GydF4y2BaB,C,D,E,F,G和H)也不是自授粉植物的单片机中所得胚胎的比例(图。GydF4y2Ba3.GydF4y2Bai,j,k,l,m和n)。此外,存在额外的GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba转基因没有能够传播GydF4y2Bap5cs2-1GydF4y2Ba突变等位基因与野生型雌蕊异花授粉的F1代GydF4y2Bap5csGydF4y2BaSesquimutant花粉(附加文件GydF4y2Ba4.GydF4y2Ba:表S1)。重要的是要注意尽管花粉表型相同(即,高频率的异常花粉颗粒,而且没有传播GydF4y2Bap5cs2-1GydF4y2Ba等位基因),NACL治疗诱导在花药中显着增加的自由脯氨酸水平GydF4y2Bap5csGydF4y2Ba昏迷的植物GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba转基因存在(图。GydF4y2Ba3.GydF4y2Baa).这些结果表明,孢子体花药组织中游离脯氨酸的水平与野生型植物相似,不足以弥补野生型植物的发育缺陷GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变体花粉。GydF4y2Ba 绒毡层或小孢子特异性启动子的选择GydF4y2Ba为了进一步研究脯氨酸合成在绒毡层(最接近花药室的孢子体细胞层)、小孢子和花粉粒中的影响,我们决定将其作为靶点GydF4y2BaP5CS2.GydF4y2Ba通过使用组织特异性启动子来这些细胞。基于可用的微阵列和文献数据,我们选择了微孔和花粉特异性启动子GydF4y2BaAT5G17340GydF4y2Ba(GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba)和绒毡层特异性GydF4y2BaLTP12.GydF4y2Ba(GydF4y2BaAT3G51590.GydF4y2Ba)启动子[GydF4y2Ba33GydF4y2Ba那GydF4y2Ba34GydF4y2Ba那GydF4y2Ba35GydF4y2Ba].这些启动子的特异性通过产生含有这些启动子的拟南芥株系来验证GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba:格斯GydF4y2Ba和GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:格斯GydF4y2Ba转录融合构建体和分析不同阶段的花药中的GUS活性模式(图。GydF4y2Ba4.GydF4y2Ba,附加文件GydF4y2Ba3.GydF4y2Ba:图S3C-F)。确认之前的观察,GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:格斯GydF4y2Ba诱导强GUS活性特别在绒毡层在阶段9和花药发育的10,并且当绒毡层退化GUS表达消失。在相同的发育阶段,GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba:格斯GydF4y2Ba在配子体发育的后期,小孢子的GUS活性更强。在花药的其他组织中几乎未检测到GUS酶活性。GydF4y2Ba 图4.GydF4y2Ba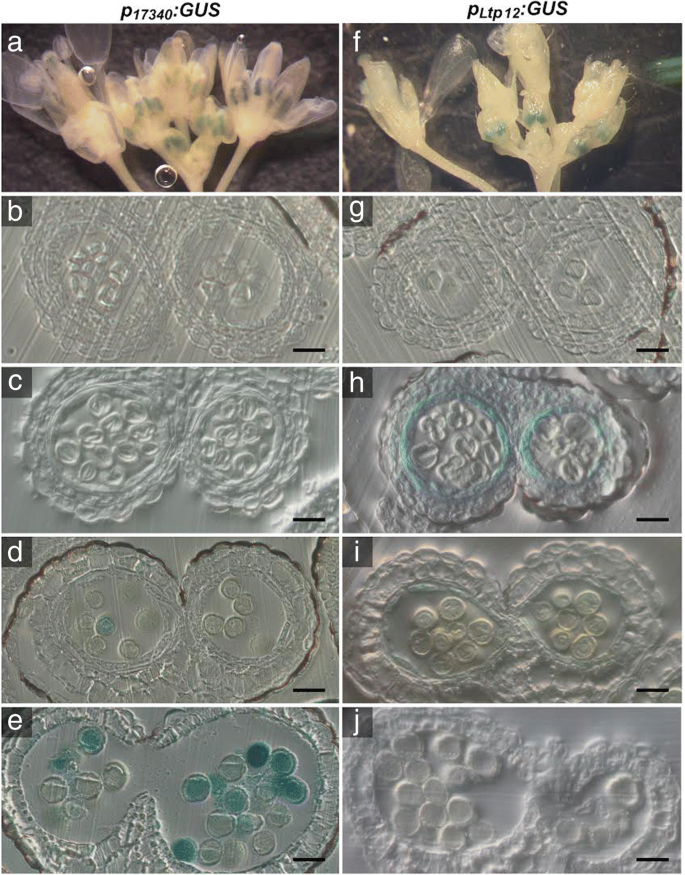
花药中Gus活性的组织化学定位GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba:格斯GydF4y2Ba和GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:格斯GydF4y2Ba转基因拟南芥植物。花序的GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba:格斯GydF4y2Ba(GydF4y2Ba安妮GydF4y2Ba) 和GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:格斯GydF4y2Ba(GydF4y2Baf jGydF4y2Ba)转基因植株用X-Gluc溶液浸润,染色过夜,固定后清除,进行显微分析。GydF4y2Ba一个fGydF4y2Ba花蕾和花在不同发育阶段的全山花序只在花药中显示GUS活性。GydF4y2Ba抵扣GydF4y2Ba和GydF4y2BaG-J.GydF4y2Ba不同发育阶段的花药横切面:GydF4y2Bab, gGydF4y2Ba第8阶段与人蛋前微孔母细胞;GydF4y2Bac、hGydF4y2Ba第9期小孢子四分体。绒毡层中有较强的GUS活性GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:格斯GydF4y2Ba(H);GydF4y2Ba维,我GydF4y2Ba第一阶段10的开始GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba:格斯GydF4y2Ba发育中的花粉表达(GydF4y2BaD.GydF4y2Ba)和花药退化绒毡层中残留的GUS活性GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:格斯GydF4y2Ba构建(GydF4y2Ba一世GydF4y2Ba);GydF4y2Bae, jGydF4y2Ba第12期花粉发育完全,GUS活性强GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba:格斯GydF4y2Ba表达式(GydF4y2BaE.GydF4y2Ba).12期花药未检测到GUS活性GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:格斯GydF4y2Ba转基因植物(GydF4y2BajGydF4y2Ba).棒= 20 μmGydF4y2Ba 全尺寸图像GydF4y2Ba Tapetum-specific表达GydF4y2BaP5CS2.GydF4y2Ba减少异常花粉的发病率,但导致生育能力恢复差GydF4y2Bap5cs1 / p5cs2GydF4y2Ba花粉GydF4y2BaTapetum细胞提供微孔发育的营养素,并有助于形成花粉外层的外层[GydF4y2Ba36GydF4y2Ba].调查if.GydF4y2BaP5CS2.GydF4y2Ba在发育中的小孢子中,绒毡层的表达可以替代脯氨酸的合成GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba构造成GydF4y2Bap5csGydF4y2BaSesquimutant植物。我们选择了超过20个主要转型体,携带两种GydF4y2Bap5cs2-1GydF4y2Ba突变等位基因和GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba其中9个携带了转基因的一个副本。在杂合子植物中,畸形花粉的比例比杂合子减少GydF4y2Bap5csGydF4y2Ba倍半植物,虽然程度不同(数据未显示)。我们选择3个纯合子系,代表中度(GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BamGydF4y2Ba)和强壮(GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BaS.GydF4y2Ba花粉发育缺陷的互补GydF4y2Bap5csGydF4y2Basesquimutant以获得更详细的分析(附加文件GydF4y2Ba4.GydF4y2Ba:表S1)。花粉中异常花粉的比例为46±3%GydF4y2Bap5csGydF4y2Ba三倍半花的数量分别为31±1%和18±2%GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BamGydF4y2Ba和GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BaS.GydF4y2Ba分别行(无花果。GydF4y2Ba5.GydF4y2Ba一种)。GydF4y2Ba 图5.GydF4y2Ba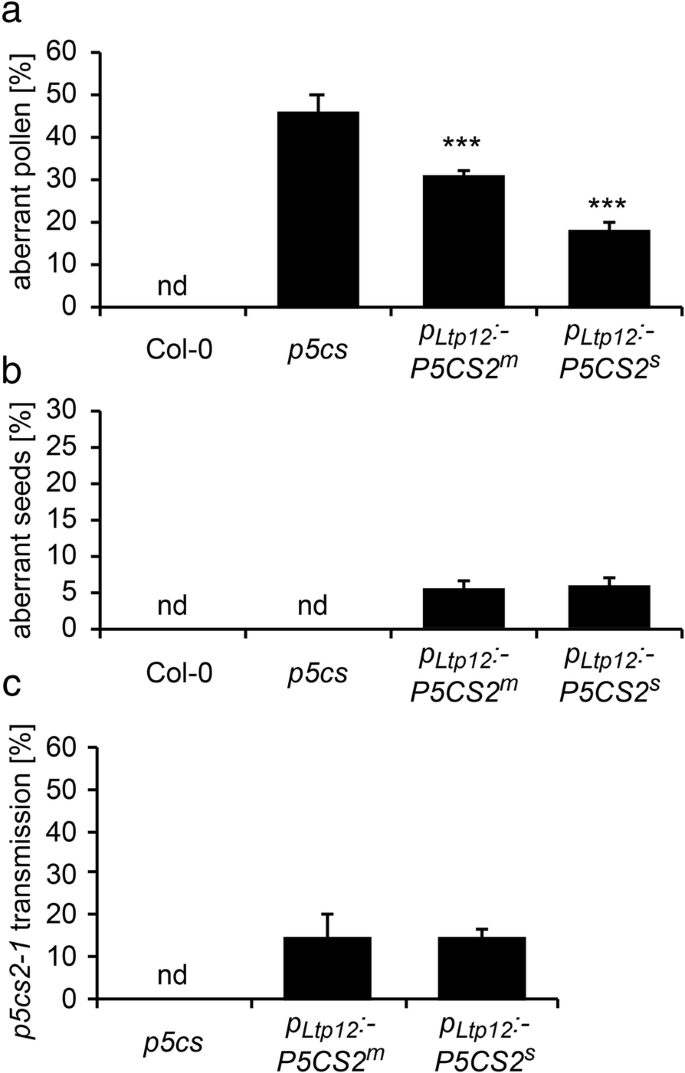
水稻花粉发育及育性分析GydF4y2Bap5csGydF4y2Ba举行的戏曲仿真GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba构造。GydF4y2Ba一种GydF4y2Ba野生型(Col-0)的异常花粉粒百分比GydF4y2Bap5csGydF4y2BaSesquimutants(GydF4y2Bap5csGydF4y2Ba) 和GydF4y2Bap5csGydF4y2Ba患有中等的癫痫症(GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba:GydF4y2BaP5CS2.GydF4y2BamGydF4y2Ba)或强烈(GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BaS.GydF4y2Ba) 表达GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba构造。nd:没有发现;***表明了显着的差异GydF4y2Bap5csGydF4y2Ba(GydF4y2BaP.GydF4y2Ba B.GydF4y2Ba在中文中描述的基因型自施剂后,SILIQUES中异常种子的百分比GydF4y2Ba一种GydF4y2Ba.GydF4y2BaCGydF4y2Ba的百分比GydF4y2Bap5cs2-1GydF4y2Ba野生型雌蕊与上述植物花粉交叉授粉获得的携带等位基因的突变苗(以抗磺胺嘧啶为标记)GydF4y2Ba一种GydF4y2Ba.酒吧GydF4y2Ba一种GydF4y2Ba那GydF4y2BaB.GydF4y2Ba和GydF4y2BaCGydF4y2Ba代表至少三个独立实验的平均值±SE。nd:未检测到。数据GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BaS.GydF4y2Ba代表来自两个独立转基因线的混合数据GydF4y2Ba 全尺寸图像GydF4y2Ba接下来,我们确定了单体中的中产胚胎的百分比GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba植物。在GydF4y2Bap5csGydF4y2Ba由于花粉谷物的不孕症,术术植物,胚胎致死的纯合体不能形成GydF4y2Bap5cs2-1GydF4y2Ba等位基因,因此是单身缺乏流入胚胎。在有效互补花粉生育率的情况下,有50%的花粉谷物GydF4y2Bap5cs2GydF4y2Ba-GydF4y2Ba1GydF4y2Ba突变会产生25%纯合GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变体,这是胚胎致命的,因为GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba在胚胎中不得授予p5cs表达。GydF4y2Ba 当。。。的时候GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba植物自交后,由于胚胎发育失败而流产的种子数量从0%增加到0%GydF4y2Bap5csGydF4y2Ba倍半植物在5±1%GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BamGydF4y2Ba6±1%GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BaS.GydF4y2Ba行(无花果。GydF4y2Ba5.GydF4y2Bab)。在用野生型雌蕊,花粉从杂交GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BamGydF4y2Ba和GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BaS.GydF4y2Ba植物传播GydF4y2Bap5cs2-1GydF4y2Ba突变等位基因到下一代的频率约为15%GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba线路远低于50%,预期的完全互补缺陷GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变花粉(图。GydF4y2Ba5.GydF4y2Bac).自交后流产胚胎的观察和传代GydF4y2Bap5cs2-1GydF4y2Ba异交花粉突变等位基因表明GydF4y2BaP5CS2.GydF4y2Ba在绒毡层表达能恢复生育能力GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变花粉,虽然只有一个相当低的程度。GydF4y2Ba 花粉特异性表达GydF4y2BaP5CS2.GydF4y2Ba完全救出异常GydF4y2Bap5cs1 / p5cs2GydF4y2Ba花粉GydF4y2Ba组织化学的GUS分析整个花序和组织学横截面的分析证实了GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba:格斯GydF4y2Ba几乎完全活跃于胃孢菌素,从拟南芥阶段开发花粉,拟南芥的阶段开发(图。GydF4y2Ba4.GydF4y2Ba,附加文件GydF4y2Ba3.GydF4y2Ba:图S3E,f)。连贯,我们生成了一个GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba构建目标GydF4y2BaP5CS2.GydF4y2Ba对发育中的花粉特异表达,并将其引入GydF4y2Bap5csGydF4y2Ba术语仿植物,以验证功能性P5CS2的花粉特异性表达是否可以拯救形态和功能性缺陷GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变体的花粉。一个有效的互补预期的结果将是在花药异常花粉数量的减少,而在长角果增加流产胚胎的数目 - 完全互补高达25%。GydF4y2Ba 我们选择了53种卡那霉素抗性转化体,其中16个携带GydF4y2Bap5cs2-1GydF4y2Ba等位基因,与观察到的非互补的传播率相似GydF4y2Bap5csGydF4y2Ba癫痫仿真。在原发性转化体的花药中,我们观察到5-20%的异常花粉晶粒,表明副本依赖性互补GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba构造。从具有单拷贝插入的转化体GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba构建后,选择4个独立的纯合子系进行进一步分析。GydF4y2Ba 在这些批次的植物中,异常花粉的比例为43±1%GydF4y2Bap5csGydF4y2Ba而在杂合子和纯合子中则分别降低到20±1%和0.15±0.10%GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba转基因植株(图GydF4y2Ba6.GydF4y2Baa, c, d, e和f,附加文件GydF4y2Ba4.GydF4y2Ba:表S1)。自授粉后GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba结果表明,在弯子叶期有胚的角果中,杂合子的败育率为12±1%,纯合子的败育率为24±2%GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba植物(图GydF4y2Ba6.GydF4y2Bab, g, h, i和j)。24±2%的缺陷胚胎在纯合子中出现GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba植物表明GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba恢复了生育能力GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变花粉但无法恢复胚胎的胚胎发育GydF4y2Bap5cs1GydF4y2Ba/GydF4y2Bap5cs2GydF4y2Ba双突变体胚胎。与野生型雌蕊异交证实为杂合子GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba突变体植株产生26±2%的可育花粉GydF4y2Bap5cs2-1GydF4y2Ba等位基因(总共182个抗钙含量为182株),当花粉供体为纯合时增加至46±1%GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba建造(94株中43株耐磺胺嘧啶植物;无花果。GydF4y2Ba7.GydF4y2Ba,附加文件GydF4y2Ba4.GydF4y2Ba:表S1)。GydF4y2Ba 图6.GydF4y2Ba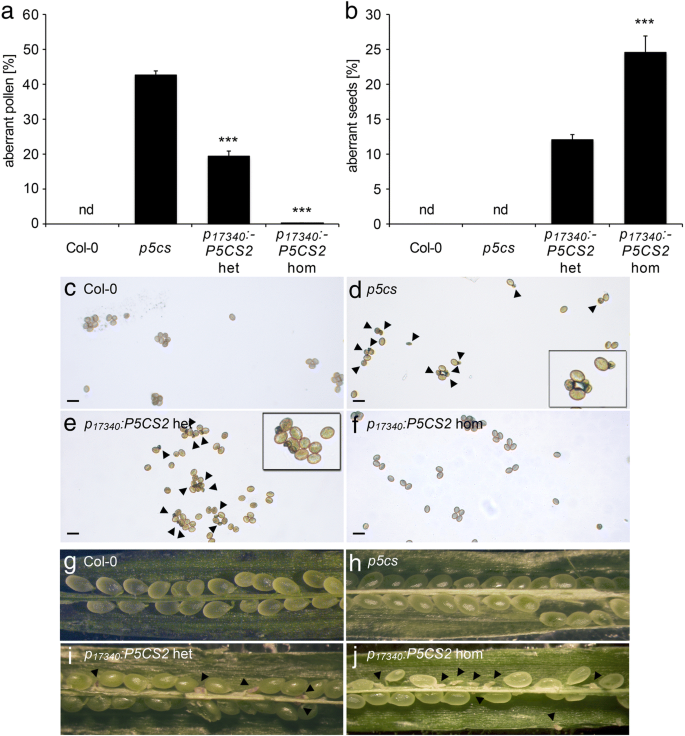
花粉和种子缺陷的形态学分析GydF4y2Bap5csGydF4y2Ba癫痫杂质补充GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba表达式。GydF4y2Ba一种GydF4y2Ba异常花粉粒的百分比和GydF4y2BaB.GydF4y2Ba野生型植物角果中异常种子的百分比(Col-0);GydF4y2Bap5csGydF4y2BaSesquimutants(GydF4y2Bap5csGydF4y2Ba)和杂合或纯合互补系(GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Bahet / hom)。柱状图代表16-22个样本的平均±SEGydF4y2Ba一种GydF4y2Ba每种基因型9个单身GydF4y2BaB.GydF4y2Ba由两个独立互补株系混合而成。nd:没有发现;在GydF4y2Ba一种GydF4y2Ba***表明了显着的差异GydF4y2Bap5csGydF4y2Ba,而在GydF4y2BaB.GydF4y2Ba***表示之间的显着差异GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Bahet,GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Bahom (p 氟GydF4y2Ba野生型花粉颗粒的明亮视野显微照片GydF4y2BaCGydF4y2Ba和GydF4y2Bap5csGydF4y2BaSesquimutantsGydF4y2BaE-F.GydF4y2Ba或者没有GydF4y2BaD.GydF4y2Ba这GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CSGydF4y2Ba建立在纯合子GydF4y2BaFGydF4y2Ba或杂合状态GydF4y2BaE.GydF4y2Ba.图中还显示了放大倍率较高的插图GydF4y2Bap5csGydF4y2Ba倍半和杂合互补系。箭头表示小而枯萎的花粉粒。GydF4y2BaG-J.GydF4y2Ba来自野生型的打开的,早产的Sysiques的立体镜像GydF4y2BaGGydF4y2Ba和GydF4y2Bap5csGydF4y2BaSesquimutantsGydF4y2BaI-J.GydF4y2Ba或者没有GydF4y2BaHGydF4y2Ba这GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CSGydF4y2Ba构造。具有致死率的异常种子GydF4y2Bap5cs2-1GydF4y2Ba纯合胚胎用箭头表示GydF4y2Ba 全尺寸图像GydF4y2Ba 图7.GydF4y2Ba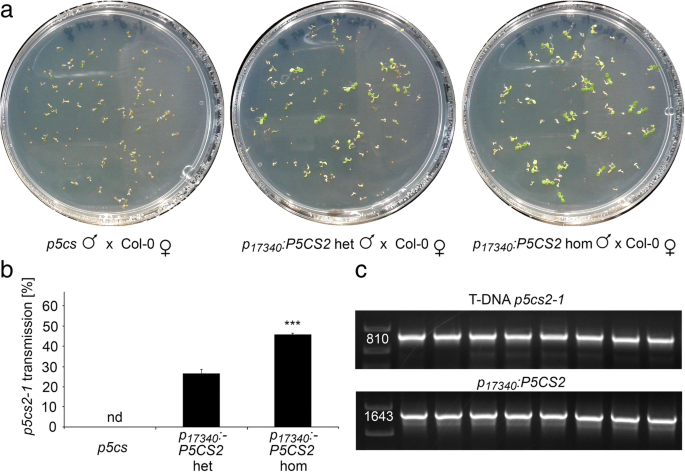
的表达GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba恢复生育力GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变体花粉。GydF4y2Ba一种GydF4y2Ba由野生型雌蕊产生的种子受精花粉从一个GydF4y2Bap5csGydF4y2BaSesquimutant(最左边的面板),或aGydF4y2Bap5csGydF4y2Ba倍半杂合子(中间面板)或纯合子(最右边面板)GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba构建物在含磺胺嘧啶的培养皿上发芽,以记录其传播GydF4y2Bap5cs2-1GydF4y2Ba等位基因。GydF4y2BaB.GydF4y2Ba如上所述,在交叉授粉后野生型雌蕊后代抗钙噻嗪抗幼苗的百分比GydF4y2Ba一种GydF4y2Ba条形图代表了至少三次独立实验的平均值±SE,实验对象为两个独立的转基因株系。nd:没有发现;***表示差异显著GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Bahet(GydF4y2BaP.GydF4y2Ba学生t检验CGydF4y2Ba磺胺嘧啶耐药后代的PCR分析。利用PCR方法对抗性植株进行分析GydF4y2BaT-DNAGydF4y2Ba生成GydF4y2Bap5cs2-1GydF4y2Ba突变(上图)和GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba构造(下半部分)。PCR产物的大小与预期碱基对数量相符(见附件)GydF4y2Ba 全尺寸图像GydF4y2Ba 花粉生育率的回收率与碱孢子中的脯氨酸浓度与花粉颗粒相关联GydF4y2Ba以验证是否有组织特异性表达GydF4y2BaP5CS2.GydF4y2Ba的确导致脯氨酸水平的改变,我们测量阶段9-10花药脯氨酸含量和成熟的野生型的花粉粒,GydF4y2Bap5csGydF4y2Ba癫痫仿生和GydF4y2Bap5csGydF4y2Ba转基因的倍半突变体GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba那GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba或GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba构造(图。GydF4y2Ba8.GydF4y2Ba一个,附加文件GydF4y2Ba4.GydF4y2Ba:表S1)。野生型花药中游离脯氨酸含量为36±2 ng/粒,野生型花粉中游离脯氨酸含量为39±2 pg/粒。的条件下GydF4y2Bap5cs2GydF4y2Ba钙质植物,脯氨酸含量约为野生型花药中含量的三分之一(12±2ng /花药)。平均通常开发和畸形的花粉,脯氨酸含量GydF4y2Bap5cs2GydF4y2Ba倍半花粉14±1 pg/粒。这两个GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba或GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba: P5CS2GydF4y2Ba对花药中脯氨酸含量有显著的促进作用,但仅对花药中脯氨酸含量有显著的促进作用GydF4y2BaP.GydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2Ba在花粉中诱导更高的脯氨酸含量。反过来,GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba对9/10期花药中脯氨酸含量无影响,但使成熟花粉中脯氨酸含量恢复到野生型花粉水平。与生育能力相比GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变花粉在不同的系,估计从胚胎致死的外观GydF4y2Bap5cs2-1GydF4y2Ba自我自行或从传输速度之后的纯合子种子GydF4y2Bap5cs2-1GydF4y2Ba等位基因与野生型雌蕊的脯氨酸含量呈较强的正相关(RGydF4y2Ba2GydF4y2Ba= 0.98,GydF4y2BaP.GydF4y2Ba 8.GydF4y2Bab),而花药中脯氨酸含量与花粉育性无关(图。GydF4y2Ba8.GydF4y2BaC)。GydF4y2Ba 图8.GydF4y2Ba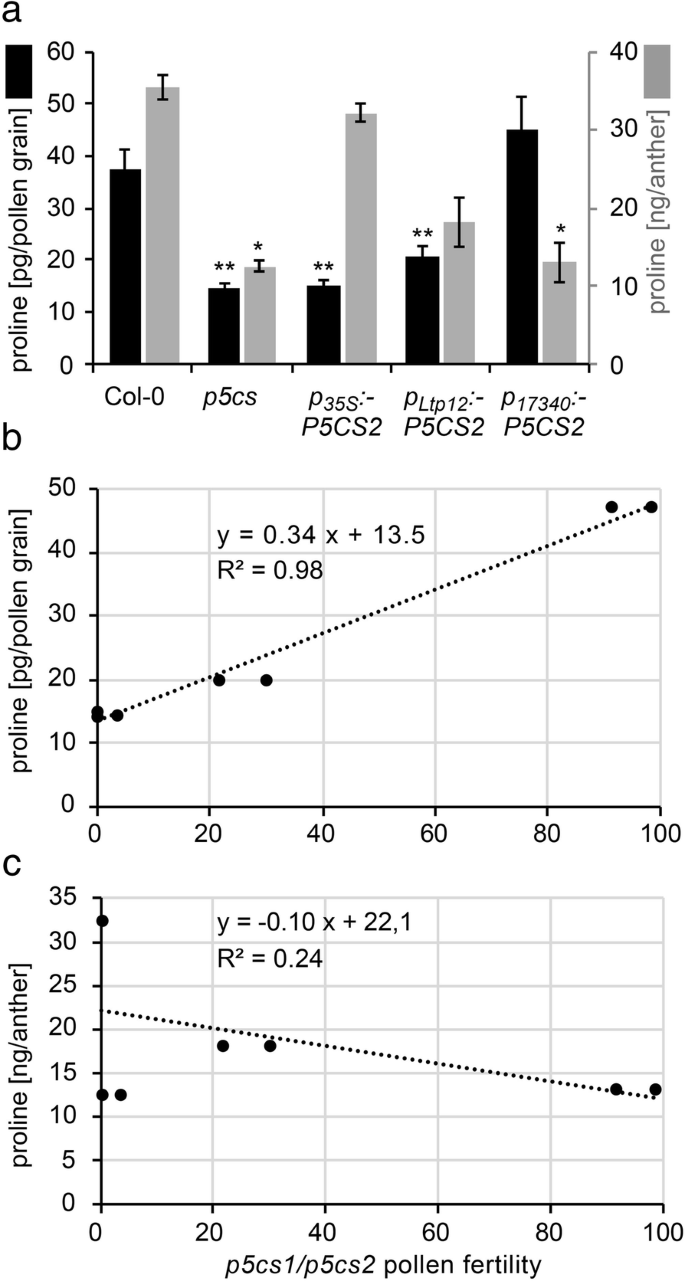
花粉育性与花粉粒中脯氨酸含量有关。GydF4y2Ba一种GydF4y2Ba花粉颗粒(黑条,左轴)和第9/10阶段的脯氨酸含量(黑条,左轴),来自野生型(Col-0)的阶段9/10波纹(灰杆,右轴),GydF4y2Bap5csGydF4y2BaSesquimutants(GydF4y2Bap5csGydF4y2Ba) 和GydF4y2Bap5csGydF4y2Ba患者携带GydF4y2BaP.GydF4y2Ba35个年代GydF4y2Ba:p5cs2,pGydF4y2BaLTP12.GydF4y2Ba: P5CS2GydF4y2BamGydF4y2Ba或者GydF4y2BaP.GydF4y2Ba17340GydF4y2Ba: P5CS2GydF4y2Ba构造。在每次分析中,平均收集了1000个花粉粒或200个9-10期的花药。条形代表两个(花药)或三个(花粉)独立样本的平均±SE。*和**表示与相应的Col-0野生型样本有显著差异(GydF4y2BaP.GydF4y2BaP.GydF4y2Bab, cGydF4y2Ba生育力GydF4y2Bap5cs1 / p5cs2GydF4y2Ba双突变花粉估计在附加文件GydF4y2Ba4.GydF4y2Ba:不同互补线中的表S1与花粉颗粒中脯氨酸的量相关GydF4y2BaB.GydF4y2Ba或9 ~ 10期花药中脯氨酸的含量GydF4y2BaCGydF4y2Ba.很强的相关性(虚线回归线)被发现在花粉粒和花粉育脯氨酸积累(R之间GydF4y2Ba2GydF4y2Ba= 0.98,GydF4y2BaP.GydF4y2Ba 全尺寸图像GydF4y2Ba |
【本文地址】