| 【科研进展】重组大肠杆菌高密度发酵的培养方式和诱导外源基因表达的研究进展 | 您所在的位置:网站首页 › 简介大肠杆菌 › 【科研进展】重组大肠杆菌高密度发酵的培养方式和诱导外源基因表达的研究进展 |
【科研进展】重组大肠杆菌高密度发酵的培养方式和诱导外源基因表达的研究进展
|
1.1.2 反馈补料法 由于非反馈补料法的补料程序是预先设定的,若微生物的生长方式与预想不同,非反馈补料法无法对其进行有效的调节,易导致乙酸生成。反馈补料法是指菌体代谢时产生的某种特殊物质(如乙酸, 二氧化碳等)会使发酵中的某些参数发生变化,依据这些参数的变化判断菌体代谢状况,再进行补料的方法。由于反馈补料法能够根据反馈的信息及时调整补料速率和策略,从而有效控制营养物质浓度,因此被广泛应用于工程菌的高密度培养。 根据反馈指标的不同,反馈补料法又可分为恒pH值补料法、恒溶解氧补料法、CER(CO2排放率)法等。常用的是恒pH 值补料法和恒溶解氧补料法。 恒pH值补料法是当营养物资充足时,菌体产酸和二氧化碳,导致培养液pH下降;当营养不足时菌体利用大量氮源产生碱性物质,导致pH值上升。因而可以把pH值的变化作为需要补料的标志,通过补充合适的补料培养基,维持培养液pH值。Jeong等采用恒pH值法反馈补料方式培养大肠杆菌,菌体密度(OD600)达到186。Kim等采用恒pH值法反馈补料培养重组大肠杆菌产人胰高血糖素样肽,菌体密度达104.2g/L。蒙健宗等在高密度发酵重组海藻糖合成酶工程菌时,采用恒pH值法反馈流加氨水和葡萄糖,维持培养液pH值为6.8,乙酸积累被控制在6.26g/L以下,补料发酵12h后获得细胞干重为51.5g/L,是分批培养的7.5倍;蛋白表达率为15.2%,是分批培养的4.5倍。但也有学者指出,由于pH值变化不完全是葡萄糖代谢的结果,该方法容易造成补料体系的错误。
恒溶解氧补料法是当营养物质充足时,菌体生长代谢旺盛,同时消耗大量的氧,导致溶解氧下降;当营养物质不足时,菌体代谢强度下降,耗氧量也减少,溶氧上升。因此可以将溶解氧的变化作为控制补料的指标,通过补充合适的补料培养基,维持培养液溶氧值在恰当水平。颜真等采用恒溶解氧法发酵工程菌DH5α/pBV-LTLTNF时,保持培养过程中的溶解氧为30%-40%,同时限制性流加葡萄糖,发酵12h后,菌体密度(OD600)达40-50,LTLTNF浓度为5.12g/L,达菌体总蛋白的50%。但胡沛臻等利用恒溶解氧补料法时发现,由于在培养初期菌体密度低,溶氧变化不明显,使用该方法易导致补料滞后。 反馈补料可以跟反馈信息及时调整补料的速率和策略,但具有一定的滞后性,不能完全避免糖浓度波动和代谢副产物的积累;非反馈补料也存在不能根据发酵环境的变化和菌体生长的具体情况作出反馈调节的缺点,因此找到一种新的、更有效的补料方法成为研究的热点和突破点。Restaino等在培养E.coli K4产荚膜多糖时,采用在发酵前期用指数补料、在发酵后期用微孔过滤的分阶段混合补料方法,取得了较好的效果,菌体浓度达到36.4g/L,多糖产量达到4.33g/L。Biener等在培养重组E.coli TB1时,建立了以菌体生长过程中产生的热量为依据进行关联补料,从而更加准确和持续控制菌体比生长速率的培养调控模型,为大肠杆菌的高密度培养提供了新的方向。 1.2 透析培养 透析培养是利用膜的半透性,根据分子不平等扩散原理使外源蛋白和培养基分离,从而除去乙酸等小分子代谢产物,解除其对生产菌的抑制作用,并维持较适宜的营养浓度,从而获得高密度菌体。Fuchs C在大肠杆菌高密度培养中通过透析培养降低乙酸含量,使细胞密度超过190g/L,蛋白浓度是分批补料发酵的3.8倍。耿艺介对不同微生物(葡萄球菌、大肠杆菌、极端环境型微生物和乳酸杆菌)的透析培养证明,细胞浓度都比其他发酵方法提高了30倍。但透析培养以生长培养基做透析液,透析过程中大量的甘油被当做废物透析掉,生产中耗资大,目前在实验室中应用较为成功,但工业化生产还有待于进一步改善。 2 诱导方式和诱导剂 高密度发酵分为生长阶段和生产阶段,生长阶段以获得较大量的菌体为目标,而生产阶段则以诱导表达外源基因从而获得大量表达产物为目标。在高密度培养中,常使用PL, Pr或Trp诱导启动子,以更好地控制目的产物的表达,避免目的蛋白积累而过早抑制细胞的生长。 为了获得高产量的表达产物,载体构建时常常插入启动子,不同启动子需要选择不同的诱导方式。目前主要有热诱导和化学诱导(IPTG诱导)两种方式。 对于热诱导,升温过程尤为重要。温度诱导型表达系统要求升温要快,因为升高温度会诱导热激蛋白的产生,从而抑制目的蛋白的表达;持续时间要短,应在2分钟内完成,因为时间太长会导致热激蛋白含量剧增,目的蛋白相对较低,为后续纯化带来困难。诱导一定时间后,由于外源蛋白的表达、代谢副产物的累积、细菌的衰老等原因,细菌会停止生长,蛋白的合成也会停止。因此要选择适宜的时机停止发酵,避免菌体自溶和目的蛋白变性。巫爱珍等人以PL启动子控制生产干扰素a-2b, 30℃生长8h后升温至42℃诱导表达2h,表达量占细胞蛋白的20%。于瑞嵩等采用30℃-42℃-40℃发酵方法,最终菌体密度OD550值为120左右。马文峰等采用2次升温诱导的方法,最终发酵的细胞密度OD600值为150,蛋白表达量为4.8g/L。Fateneh等在42℃诱导30min后再降到37℃诱导4h,取得了很好的效果。 目前,常采用的化学诱导是乳糖基因(lac)及其衍生的启动子及其融合产物( Tac , Pac , Rac ),再加入异丙基βD硫代半乳糖苷(IPTG)诱导外源基因的表达,其用量会影响表达水平。Sriubolmas等指出IPTG浓度在0.0025-0.1mmol/L时,增加IPTG浓度可提高重组青霉素G酰化酶活性,而IPTG浓度在0.2-0.5 mmol/L时,酶活性下降。胡沛臻等构建人黑色素瘤抗原MAGE1/HSP70/MAGE3融合蛋白基因重组工程菌,对数生长期时用200μmol/L IPTG 诱导5h,菌体量提高至70g/L以上,目的蛋白表达量占菌体蛋白总量的38%以上。YILDIRIM S等研究表明,IPTG浓度为0.5mmol/L,是细菌生长和产物表达的最佳浓度;何勇智等采用0.5mmol/L IPTG进行诱导,发酵产量比传统的LB培养基及培养方法提高了6倍多。尽管IPTG的诱导效果令人满意,但IPTG 价格昂贵,造成生产成本增加,而且毒性对人体产生严重伤害,对发酵所得的生物产品带来不利影响,因此它在大规模发酵生产中的应用受到了限制。为了寻找ITPG的替代物,国内外学者做了大量的尝试。有人尝试用乳糖代替IPTG,并取得了不错的效果。陈亮等在用乳糖诱导重组大肠杆菌BL21表达三苯基甲烷类染料脱色酶TpmD时发现,目的蛋白表达量占总蛋白量的36.62%,与IPTG的诱导结果(35.03%)无明显差别,而且收获的菌体量比用IPTG诱导时更高。WU PH等通过乳糖诱导的分批补料培养,获得的全细胞生物催化剂的生产效率比IPTG诱导的分批培养提高了6倍。卫红飞等报道,用4g/L乳糖诱导工程菌4h,得到重组蛋白表达量与0.1% IPTG 诱导工程菌3h 后相似,说明适当延长乳糖诱导时间,可以得到较高的表达量。吴一凡等在诱导重组大肠杆菌表达B淋巴细胞刺激因子时综合比较了乳糖和IPTG的诱导效果,发现用乳糖作为诱导剂时,目的产物占总蛋白量的15.7%, 略低于IPTG的诱导效果,但成本仅为用IPTG诱导时的3%。李兆鹏等发现,乳糖的诱导表达量只能达到IPTG的55%。尽管乳糖的诱导效果不及IPTG,诱导过程也比IPTG 复杂的多,但乳糖无毒、价廉,且其本身又可作为一种碳源被大肠杆菌利用,因此乳糖替代IPTG作为诱导剂的研究对工业化发酵生产重组蛋白具有重要的意义。 重组大肠杆菌高密度高表达发酵是基因工程产业化的关键,也是基因工程技术从实验室通向市场的桥梁,改进高密度发酵培养方式、优化诱导方式和诱导剂,对实现重组大肠杆菌高密度高表达发酵工业化生产具有重大意义。 天津威特 宗惠 编审 李永霞
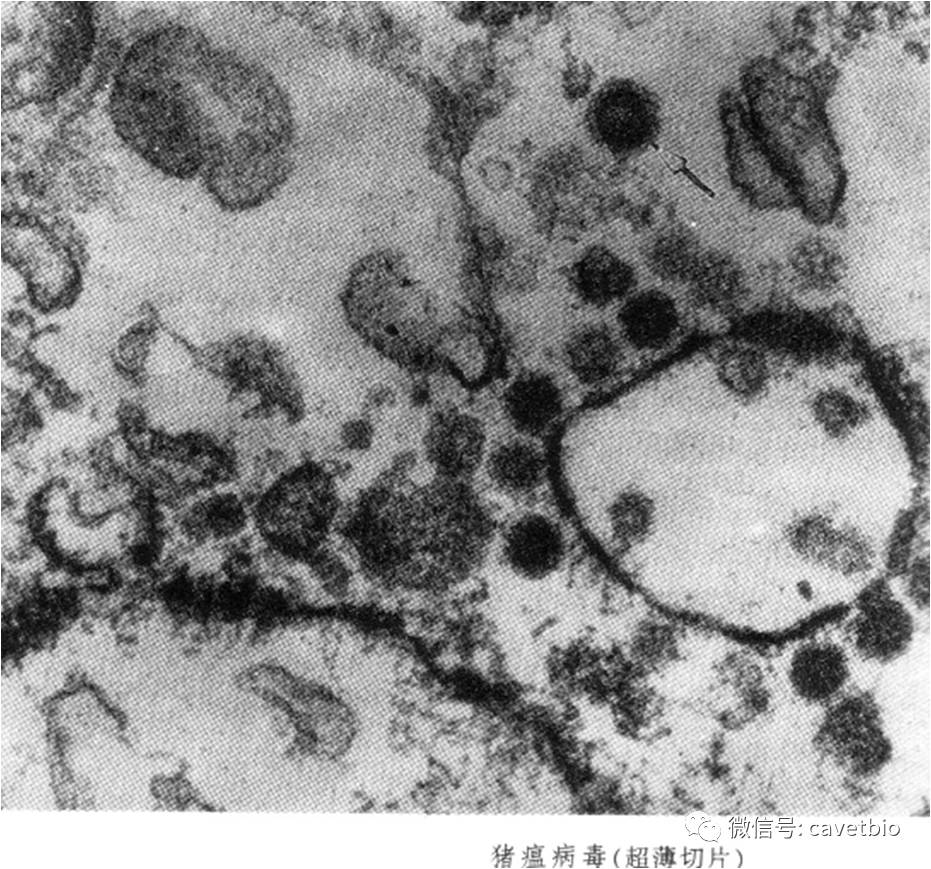
猪瘟的研究进展 猪瘟(CSF),俗称“肠烂瘟”,是全球范围最重要的流行性病毒性疾病之一。CSF病毒(CSFV)是引起该病的致病因子。CSFV为有囊膜的单股正链RNA病毒。基于病毒基因的部分序列分析,可以将CSF分为三种基因型,但是,与病毒的毒力无直接相关性。由于病毒和宿主因素,猪瘟的临床症状表现出多种形式,因此,除了仔细观察剖检变化之外,实验室多种诊断方法是必不可少的。猪对猪瘟极少有天然免疫力,因此,研究猪瘟的免疫方法是预防猪瘟的基础工作。本文,我们将从病原学,流行病学,临床症状与病理变化,发病机理与免疫效应,疾病诊断和免疫防制方面进行简单介绍。 1. 病原学 1.1 病毒属性 猪瘟病毒(CSFV)属于黄病毒科、瘟病毒属,由大约12.3kb的单股正链RNA基因组构成,通过病毒和细胞蛋白酶对前体蛋白翻译处理得到13种成熟的蛋白质,包括4种结构蛋白(核衣壳蛋白C,囊膜蛋白E1,E2和糖蛋白Erns)和9种非结构蛋白(Npro, p7,NS2-3,NS2,NS3,NS4A,NS4B,NS5A和NS5B)。后者在病毒复制中具有各种功能,例如,NS5B可以替代RNA依赖性RNA聚合酶,NS3具有类似蛋白酶的作用等。 1.2 病毒理化特性 CSFV在不同环境条件下的存活差异很大,尤其是在受到温度影响的条件下。一般来说,CSFV在寒冷,潮湿及富含蛋白质的条件下有很长的存活时间。排泄物中的CSFV,在5℃的条件下能存活几天到数周,如果温度升高到35℃,CSFV的存活时间大大缩短。研究显示,CSFV在60℃ 3min的实验条件下即可立即被灭活,对于污染的猪圈,CSFV可以存活几天,在寒冷的冬季条件下甚至可以存活一个月。关于pH值,CSFV在pH5和pH10之间是相对稳定的。低pH值下,CSFV的半衰期与温度相关,室温下CSFV的平均半衰期比在4℃下低至少10倍(4℃时为70小时,pH为3时为21℃5小时)。 2 流行病学 2.1 易感动物 目前,猪科的不同成员都可作为易感宿主,尤其是家猪和欧洲野猪。另外,最近发现普通疣猪和非洲灌丛野猪也有易感性。该病的易感性与猪的性别,年龄,营养状况无明显关系。 2.2 传染源 病猪是最主要的传染源,急性型病猪全身带毒,其中脾脏和淋巴结等免疫器官含毒量最高。另外,感染本病后康复的猪,处于潜伏期的猪和隐性感染的猪均携带该病毒,可作为本病的传染源。被猪瘟病毒污染的饲料,饮水,饲养工具等也可大量散播病毒,造成猪瘟的发生和流行。 2.3 传播途径 传播途径分为水平传播和垂直传播两种形式。水平传播途径是病毒通过消化道粘膜、呼吸道粘膜、生殖道粘膜、眼结膜或擦伤的皮肤等途径直接进入易感动物组织内传播感染;垂直传播主要指患病母体通过胎盘将病毒传至子代,当母猪怀孕期间发生感染时,病毒能通过胎盘屏障感染子宫内的胎儿,这有可能导致仔猪持续感染。虽然母猪常常仅显示轻微的临床症状,但依赖于妊娠期的感染能导致初生胎儿木乃伊化,流产或死胎。在妊娠第50~70天感染的母猪,可引起免疫耐受现象而使后代出生后持续感染。
本病无明显季节性,一年四季均可发病,其中春秋两季极为明显。初次感染猪瘟的猪群,主要以急性型经过,发病率与死亡率在90%以上。 3 临床症状与病理变化 猪瘟根据病程时间大致可以分为以下几种型:急性型,慢性型和持续型。急性型一般在感染CSFV 2~4周死亡。死亡率可达100%。在感染的前两周,临床症状主要表现为高热,厌食,胃肠症状,全身无力和结膜炎。感染后二至四周可能发生神经系统症状,包括不协调,麻痹,麻痹和抽搐,最后多以死亡告终。期间主要出现典型的败血症变化,皮肤、粘膜、浆膜和实质器官可见大小不一的出血样病变。慢性型多发生在猪瘟流行地区。病猪多发生间歇性发热,抑郁,消瘦和弥漫性皮炎。长期感染的动物常成为僵猪,最后死亡。本型特征性病理变化为大肠轮层状溃疡,俗称“扣状肿”。持续型感染猪常作为病毒携带者,无明显病理症状,无热或仅出现轻热,偶有病猪出现皮肤坏死或行走不稳等轻微病症,病死率不高。 4 发病机理与免疫效应 综上所述,根据CSFV病毒毒株的毒力和宿主因素不同,病猪的临床症状可从急性致死型到无明显症状的持续型不一。其中非特异性临床症状占主要地位,并且,与其他几种猪的传染病的鉴别只能通过实验室诊断。急性致死型多伴随病毒性出血症,严重的血小板减少,肺水肿,淤血和出血。有实验证明,急性脑脊液中可见许多病变细胞因子。 CSFV侵入机体后首先在扁桃体中进行复制,随后扩散至周围淋巴组织。病毒通过淋巴管到达区域淋巴结。病毒通过血液传播到次级复制部位,如脾脏,骨髓和内脏淋巴结,在这里进一步复制。在几个巨噬细胞群体中可观察到凋亡反应以及吞噬和分泌激活。这些活化的巨噬细胞似乎在(免疫)发病机制中发挥关键作用。此外,干扰素(IFN)系统的干扰对发病机制有所贡献。血清中IFN-α与疾病的严重程度和毒性的相关性似乎存在相关性。在早期感染后两天,临床症状发作之前可检测到高浓度的IFN-α。 5 疾病诊断 快速可靠的诊断对于及时实施针对CSF的防控措施至关重要。相关的实验室诊断方法以及取样和运输指南可参考世界动物卫生组织(OIE)的《Manual of Diagnostic Tests and Vaccines for Terrestrial Animals》(《陆生动物诊断试验和疫苗手册》)和《EU Diagnostic Manual》(《欧盟诊断手册》)。 下面介绍几种常见的实验室诊断方法: ①病毒分离:CSFV可以再在不同的永久细胞系上分离培养,如猪肾细胞系PK15或SK6。在PK15中增值的病毒可用聚乙二醇(PEG)及硫酸铵沉淀后,再经超速离心及梯度离心后,得到纯化的病毒。 ②CSFV基因组检测:RT-PCR方法是检测CSFV最基本也是最可靠的方法之一,PCR扩增的产物可以直接用来测序,进行毒株的分型。其敏感性高于病的分离,但缺点在于不适用于大批量检测。 ③ELISA抗体检测: ELISA是目前最常用的CSFV抗体检测方法之一。现有的针对结构蛋白的抗体检测使用最广泛的是CSFV E2 结构蛋白的抗体检测,具有极高的敏感性和特异性。 6 免疫防制 猪对猪瘟病毒极少有天然免疫力,只有在感染后或者免疫后才能得到保护。然而,本病的感染率和致死率极高,极少有猪能在感染后健康存活。因此,免疫接种是目前猪瘟最可靠的防治措施。 种公猪实行每年间隔免疫2次;后备母猪在配种前7~10天进行肌肉免疫;生产母猪在产后25天进行肌肉免疫;对于新生仔猪,如果母源抗体水平良好,且当地无猪瘟疫情,可以在产后25天进行猪瘟疫苗的免疫,进行肌肉注射;如果当地及周边地区猪瘟疫情发展比较严重,应在出生后立即免疫,2小时后再喂奶。发挥主动免疫和被动免疫双重保护效果。在进行猪瘟疫苗免疫注射时,要做到以下几点:在疫苗使用前2天和后3天严禁使用抗病毒药、具有免疫抑制性的抗生素类药物及消毒药液,猪瘟疫苗要与猪繁殖和呼吸综合征疫苗间隔15d以上使用,与其他疫苗间隔时间为7~10 d。在不使用疫苗期间,可做好消毒药的使用工作,一般规模猪场为一周1~2次,做好消毒防病工作。 若爆发紧急疫情,及时上报相关部门,积极配合工作。隔离疑似感染猪群,将病猪群进行无害化处理,对健康猪群进行加强免疫。建议猪场每天消毒1~2次。 销售公司 刘俊 编审 李克斌返回搜狐,查看更多 |

【本文地址】