| 【学术笔记】第十三篇:植物的光周期现象和季节性 | 您所在的位置:网站首页 › 植物开花控制的途径是什么 › 【学术笔记】第十三篇:植物的光周期现象和季节性 |
【学术笔记】第十三篇:植物的光周期现象和季节性
|
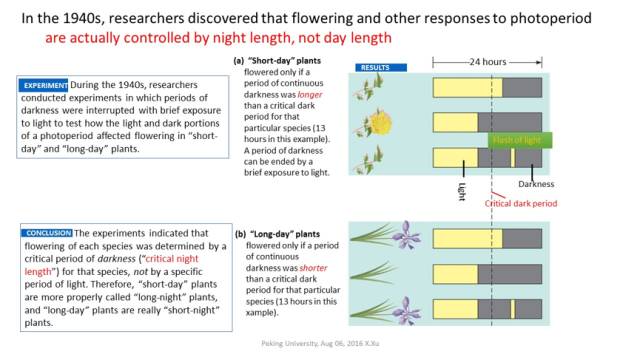
图1. “Maryland Mammoth”烟草与光周期现象 在短日照条件下的夜间进行短时间光照后,短日照植物不会开花;而长日照植物在夜晚长度大于开花临界值时,在夜间给与短暂的光照处理会促进其开花。这表明植物的光周期现象依赖夜间的长度(Critical Night-Length, CNL),而不是白天的长度(图2)。
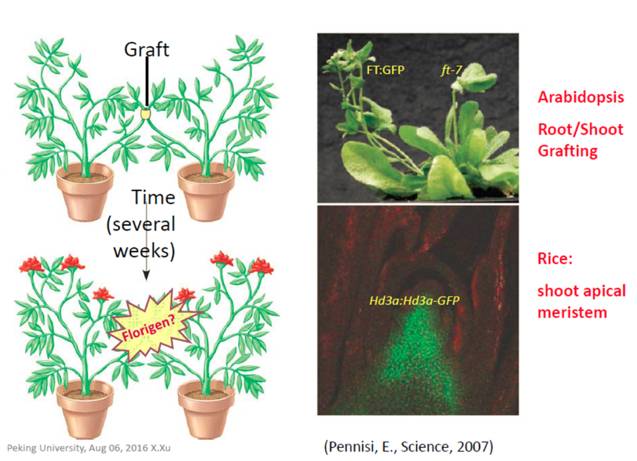
图2. 植物光周期现象(如开花)取决于夜间的长度 在二十世纪30年代,前苏联科学家Mikhail Chailakhyan发现,生长在非适合自身开花光周期条件下的植物与开花植物嫁接或利用开花植物叶片提取物处理后,均可促成其开花,他将这种可能在叶片合成运输至芽尖促进开花的物质称之为“成花素(Florigen)”。赤霉素(GA)可以促进部分长日照植物在短日照条件下开花,但不能促进短日照植物在长日照条件下开花,不完全符合成花素的特性;韧皮部短时间内蔗糖浓度的增加同样可以促进花的形成,但只能在强光照条件下检测到蔗糖浓度的短时间上升,并没有在弱红光或远红光条件下检测到,表明蔗糖不是促进植物开花所必需的因子。在70年之后,研究人员发现在韧皮部表达的球状蛋白FLOWERING LOCUS T (FT) 可以运输至芽尖促进开花,能满足成花素的特性(图3)。
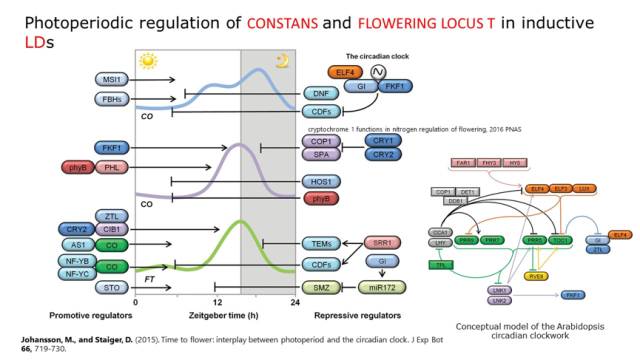
图3. 成花素Florigen与FLOWERING LOCUS T 光周期调控的开花途径受内源生物钟和外源光信号的直接调节,CONSTANS(CO)是调控 FT表达的重要转录因子,CO可结合 FT启动子区的CORE顺式作用元件促进其表达, CO的表达受生物钟和光信号通路的共同调控。在长日照条件下,GI-FKF1复合体通过促进CDF1/2的降解,增强 CO的表达;ELF4通过调控GI的亚细胞定位抑制 CO的表达。红光受体phyB可能与HOS1互作在清晨促进CO的降解;蓝光受体cry1/2通过抑制COP1-SPA复合体的活性促进CO在傍晚的积累(图4)。最近的研究表明,TOC1和PIF3可形成复合体调控短日照条件下靶基因的表达和植物的生长发育。
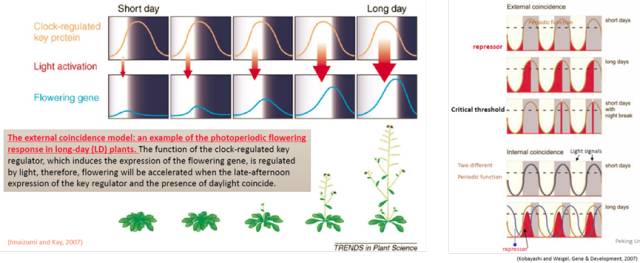
图4. 生物钟和光信号共同调控CO/FT的表达 长日照植物开花模型为,一个促进开花基因的表达受生物钟组分和光信号的共同诱导,当日照时间足够长时,开花基因的表达才能达到足够的丰度,促进开花。短日照植物开花有外源(External coincidence)和内源(Internal coincidence)两个模型,外源模型为开花抑制因子表达的模式是固定的,但其功能依赖与光照,长日照条件下其表达峰值时有光照,可抑制开花;而内源模型展示了一个开花抑制因子与其抑制子功能随光周期变化的动态过程,在短日照条件下两者相位一致,开花抑制因子的活性被抑制,而在长日照条件下两者相位不一致,开花被抑制(图5)。
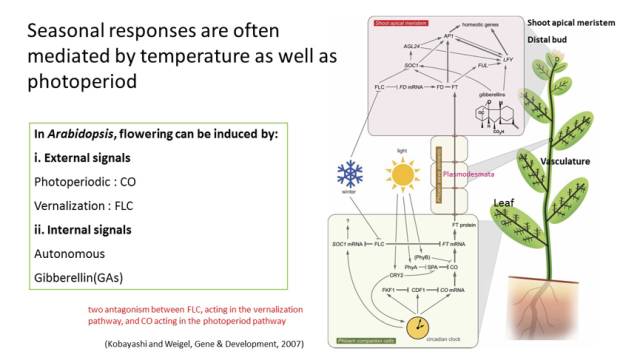
图5. 长日照或短日照条件下开花时间调控模型 在自然条件下,光周期现象与温度相关联,比如一些高纬度或高海拔地区的植物必需经历一个低温的春化处理,才会在春天开花。被低温抑制的FLC可以通过不依赖于CO的途径调控开花,这个过程被称之为春化途径;同时,FLC通过抑制CO对FT的激活作用抑制开花(图6)。高温促进拟南芥开花的过程与光周期调控开花之间也存在联系,CO和PIF4/5参与短日照条件下高温对植物开花的促进过程。生物钟通过整合光、温度等环境信号,调控植物自身生长发育过程与环境周期性变化的同步化,增强植物生存和环境适应性。
图6. 光周期、温度调控与季节性响应 Reference Garner, WW, Allard, HA (1920). Effect of the relative length of day and night and other factors of the environment on growth and reproduction in plants. J. Agric. Res. 18, 553-606. Kobayashi, Y., and Weigel, D. (2007). Move on up, it's time for change--mobile signals controlling photoperiod-dependent flowering. Genes Dev 21, 2371-2384. Krzymuski, M., Andres, F., Cagnola, J.I., Jang, S., Yanovsky, M.J., Coupland, G., and Casal, J.J. (2015). The dynamics of FLOWERING LOCUS T expression encodes long-day information. Plant J 83, 952-961. Johansson, M., and Staiger, D. (2015). Time to flower: interplay between photoperiod and the circadian clock. J Exp Bot 66, 719-730. Soy, J., Leivar, P., Gonzalez-Schain, N., Martin, G., Diaz, C., Sentandreu, M., Al-Sady, B., Quail, P.H., and Monte, E. (2016). Molecular convergence of clock and photosensory pathways through PIF3-TOC1 interaction and co-occupancy of target promoters. Proc Natl Acad Sci U S A 113, 4870-4875. Fernandez, V., Takahashi, Y., Le Gourrierec, J., and Coupland, G. (2016). Photoperiodic and thermosensory pathways interact through CONSTANS to promote flowering at high temperature under short days. Plant J 86, 426-440. Corbesier, L., Vincent, C., Jang, S., Fornara, F., Fan, Q., Searle, I., Giakountis, A., Farrona, S., Gissot, L., Turnbull, C., and Coupland, G. (2007). FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 316, 1030-1033. Ibanez, C., Kozarewa, I., Johansson, M., Ogren, E., Rohde, A., and Eriksson, M.E. (2010). Circadian clock components regulate entry and affect exit of seasonal dormancy as well as winter hardiness in Populus trees. Plant Physiol 153, 1823-1833.
|
 返回搜狐,查看更多
返回搜狐,查看更多【本文地址】