| 徐鹏报告详细解读:鲤鱼异源四倍体起源及基因组进化 | 您所在的位置:网站首页 › 斑马鱼染色体特点 › 徐鹏报告详细解读:鲤鱼异源四倍体起源及基因组进化 |
徐鹏报告详细解读:鲤鱼异源四倍体起源及基因组进化
|
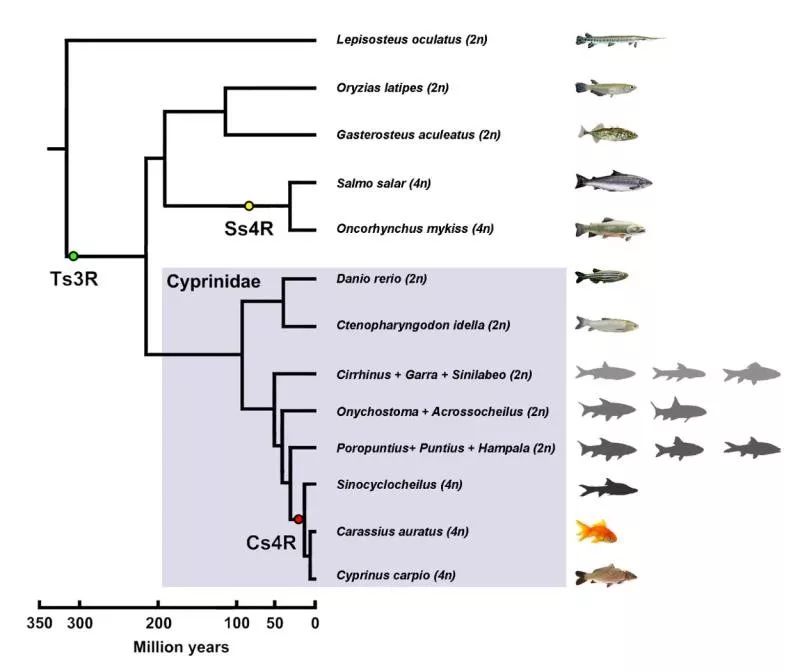
异源四倍体 - 鲤 作为世界上最重要的淡水养殖鱼类之一, 鲤(cyprinus carpio)共有100条染色体,是一种异源四倍体硬骨鱼类。鲤除经历了硬骨鱼类共有的第三轮全基因组复制事件(Ts3R)外,还经历了特有的第四轮全基因组复制事件(Cs4R),因此也是广受关注的研究脊椎动物多倍体基因组演化的理想模式生物。
基因组倍增是生命演化过程中重要的演化驱动力 | 正文图1 早在1967年,演化生物学界泰斗Susumu Ohno先生就已发现,鲤染色体数目是其他大多数鲤科鱼类的2倍,并且其减数分裂中是形成50个二价体,而非25个四价体。在其1970年出版的专著《Evolution by Gene Duplication》中,Susumu Ohno先生提出“基因组复制驱动进化”的理论。自此,鲤异源四倍化现象进一步受到科学界的关注,各国学者纷纷从分子遗传学层次开展了大量有关鲤四倍化起源的研究。
Susumu Ohno & 《Evolution by Gene Duplication》 近年来,测序技术的发展使得利用全基因组尺度的遗传工具更全面揭示鲤四倍化形成机制和全基因组复制后基因的演化成为可能。 2014年,在课题组发表于Nature Genetics的研究中,我们通过对松浦镜鲤与斑马鱼进行比较基因组分析,发现鲤和斑马鱼的基因组呈现出明显的“2:1”的染色体共线性关系,注释获得的鲤功能基因的数量也接近二倍体鲤科鱼类的两倍,这为我们后续有关鲤第四轮基因组复制和异源四倍体起源的研究提供了组学证据。 鲤异源四倍体的祖先溯源 然而,关于鲤的身世,仍有太多未解之谜。
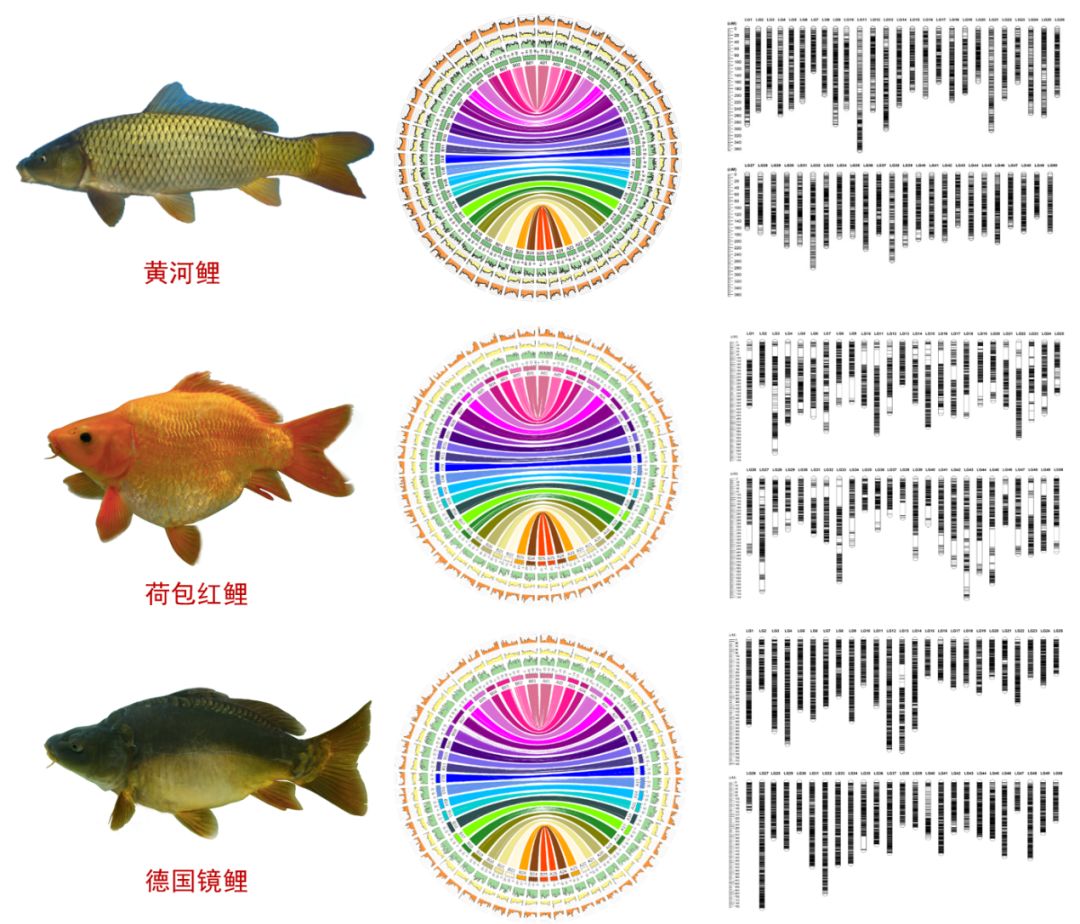
为了回答上述问题,我们开展了一系列研究: 1 数据储备 要解开以上这些问题的答案,首先需要更翔实的基因组数据作为支撑。由此,我们开展了鲤3个亚种(黄河鲤、荷包红鲤、德国镜鲤)的全基因组测序工作,获得了三个染色体水平的高质量鲤参考基因组。并基于鲤250K SNP分型芯片,构建了3个高密度遗传连锁图谱。
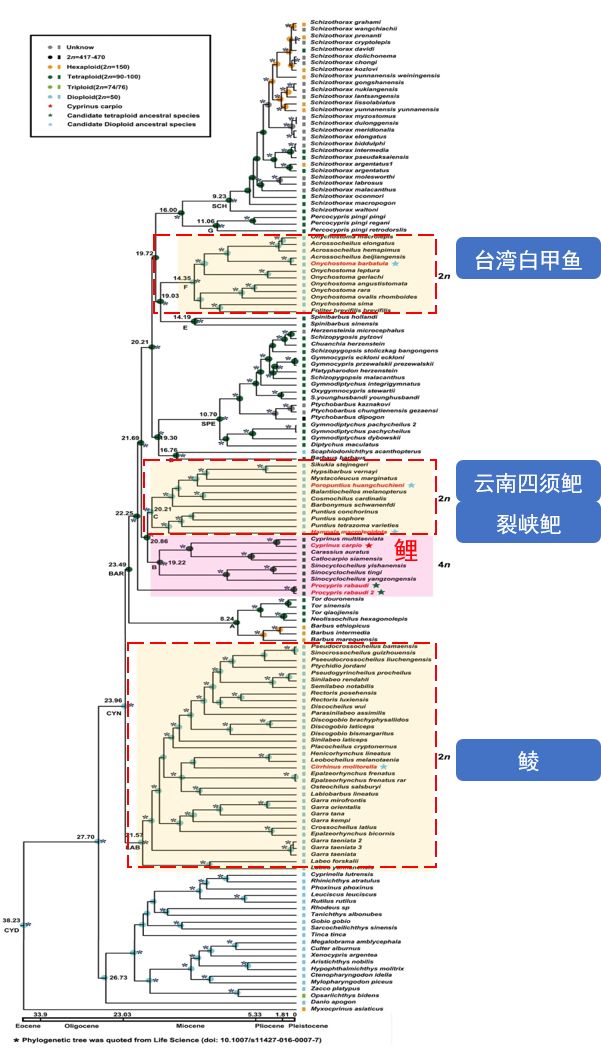
鲤基因组物理图谱、遗传连锁图谱 | 附图1、2 & 4 2 祖先溯源 接下来,我们要回答的是“鲤的祖先是谁?” 鲤亚科可分为4个类群,分别为鲃类(barbine)、鲤类(cyprinine)、野鲮类(labeonine)和裂腹鱼类(schizothoracine)。2016年,中科院水生所何舜平研究员团队在《中国科学》期刊上发表了关于鲤亚科多倍体物种独立起源的相关研究论文,为异源四倍体鲤的二倍体祖先溯源工作提供了重要线索。他们分析了鲤亚科46属123种鱼类以及20种鲤亚科以外的其他鲤科鱼类,利用线粒体和核基因序列构建了系统发育关系(下图),并以此来追溯裂腹鱼类多倍体的起源及其物种分化。
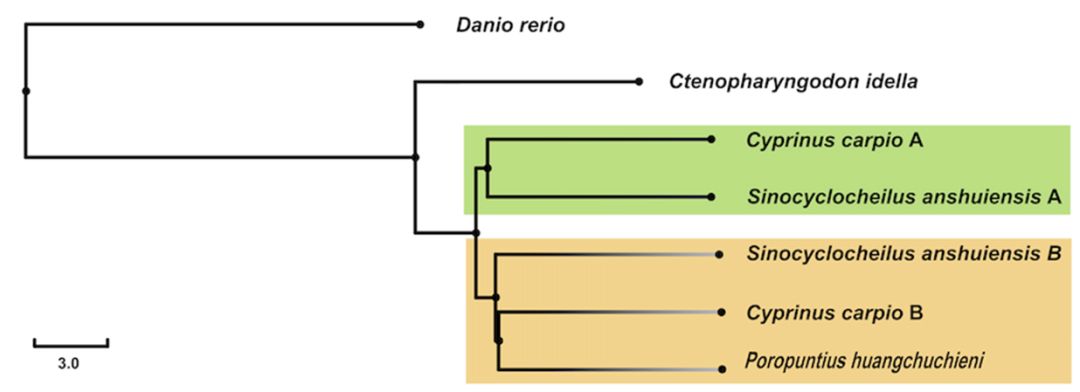
鲤亚科系统发育关系 | 改绘自参考文献2 在这张精细的鲤亚科系统发育树上,可以识别出与异源四倍体鲤的3个近缘二倍体类群,为我们寻找可能的鲤的祖先二倍体提供了重要线索。基于此,为探讨鲤及其近缘四倍体和二倍体鲤亚科物种的进化关系,我们利用核基因重组激活基因2(rag2)重构了鲤鱼亚科代表种的系统发育树(下图左),发现该基因在二倍体鲤科鱼类中仅存在一个拷贝,但在四倍体鲤中存在两个拷贝。
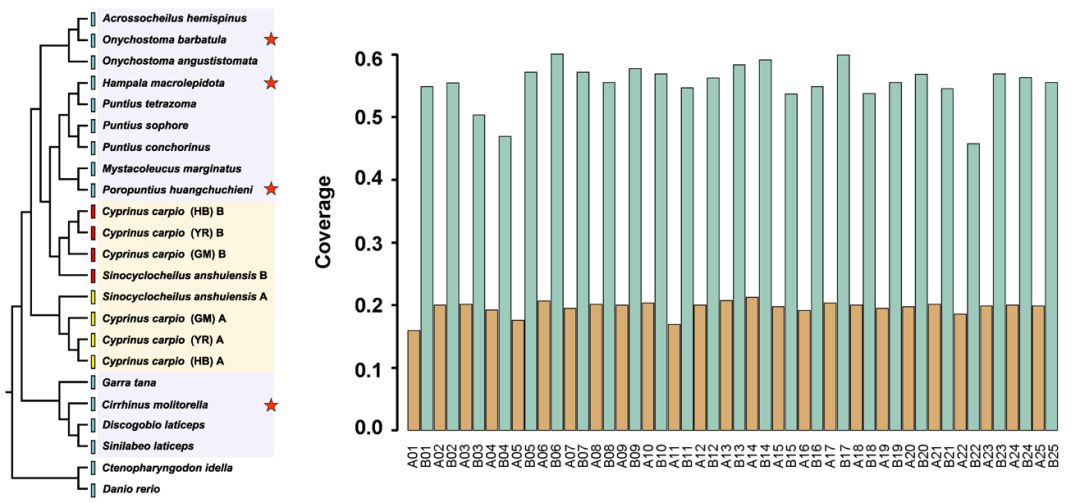
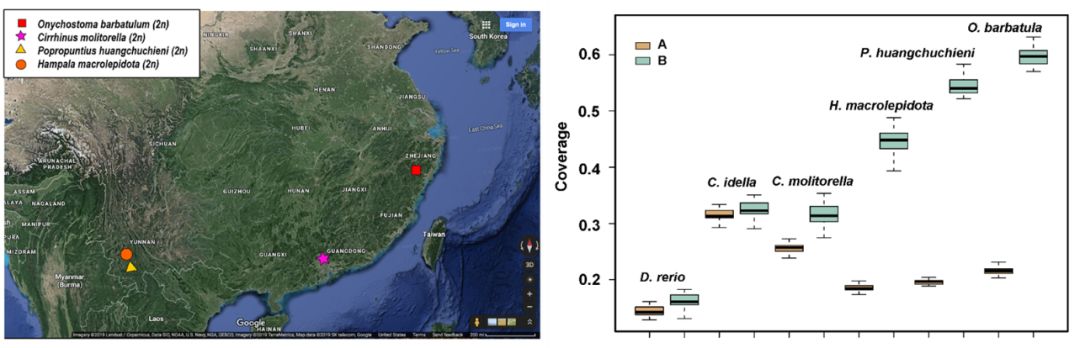
鲤亚科代表性物种系统进化关系 & 四须鲃基因组与鲤基因组的覆盖度 | 正文图2a & 2b 从系统进化树可以看出,鲤的其中一个rag2 基因与鲃系聚为一支,提示其两套亚基因组中的一个可能来源于鲃系的二倍体物种。而另一个rag2 基因则单独聚为一支(正文图2a)。 受到这样的系统发育关系的启发,我们猜想,是否可以借助鲤近缘二倍体物种的基因组序列信息将其两套亚基因组进行拆分? 由此,我们选取了与鲤亲缘关系较近的云南四须鲃(Poropuntius huangchuchieni)进行全基因组测序。通过与鲤基因组序列的比对,我们发现,四须鲃基因组序列与鲤的同源染色体对(Homeologous chromosome pairs)中的一条染色体相比具有较高覆盖度(genome coverage),而与另一条染色体相比覆盖度则较低,并且这种现像在25对同源染色体中均存在。借此,我们成功将异源四倍体鲤的基因组拆分为两套来自祖先二倍体的亚基因组,各由25条染色体组成,分别命名为A亚基因组和B亚基因组。 为了更加接近鲤身世之谜的真相,我们又选取了鲃亚科另外两个近缘二倍体类群中的台湾白甲鱼(Onychostoma barbatulum)和裂峡鲃(Hampala macrolepidota),以及亲缘关系稍远的二倍体类群野鲮亚科中的鲮(Cirrhinus molitorella)进行全基因组测序,并以基因组已公开的斑马鱼(Danio rerio)和草鱼(Ctenopharyngodon idella)作为参考序列进行比较基因组学分析。 3个鲃亚科物种与异源四倍体鲤的比较基因组学分析结果显示,三者与B亚基因组比对的基因组覆盖度均显著高于A亚基因组,而草鱼(鲤科雅罗鱼亚科)、斑马鱼(鲤科
亚科)与鲤两个亚基因组相比,基因组覆盖度差异则不大(正文图2c)。
近缘二倍体物种采样地点及与鲤基因组覆盖度 | 附图6 & 正文图2c 综合上述基于核基因序列的鲤亚科分子系统学研究,以及多个二倍体鲤科鱼类全基因组测序和比较基因组分析,我们鉴定出“吻孔鲃Poropuntius-小鲃Puntius-裂峡鲃Hampala”为B亚基因组最可能的祖先二倍体类群之一,而A亚基因组的祖先可能为鲤科中亲缘关系相对较远的未知物种,甚至可能是已经灭绝的物种。 为了验证这种系统进化关系,我们从单个rag2基因延伸至鲤:金线鲃:斑马鱼/草鱼/四须鲃 为2:2:1关系的2,071保守同源基因,并利用这些基因分别构建系统进化树得到整合结果,对鲤的亚基因组起源进行佐证。
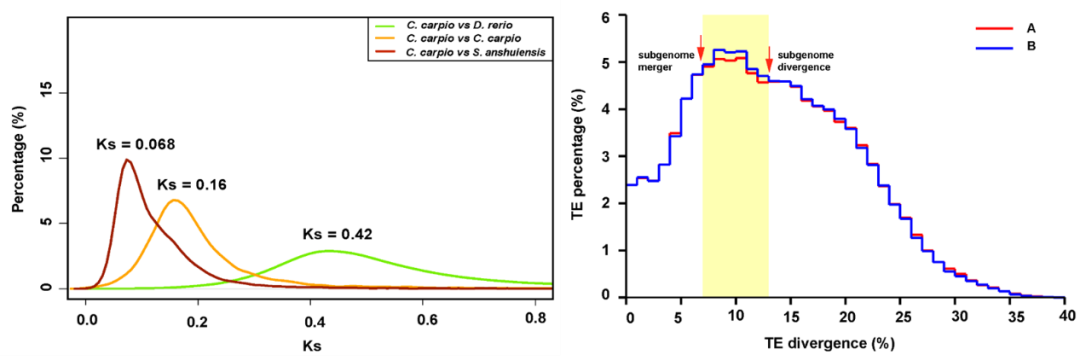
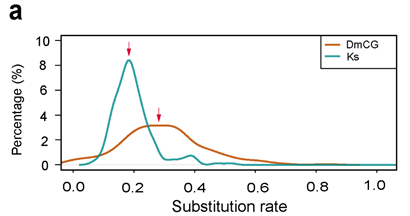
2,071个保守同源基因佐证鲤的亚基因组起源 | 附图7 3 鲤异源四倍化形成时间推演 什么时候形成了异源四倍体鲤? 为了计算鲤异源四倍化发生时间,利用斑马鱼、金线鲃、鲤1:2:2关系的8,270对同源基因计算得到的同义替换率Ks和已有的鲤分子钟3.51x10-9,推算出鲤和斑马鱼的分化时间为6000 万年前(60 Mya),鲤A、B亚基因组分化时间约为2300万年前(23 Mya)。而亲缘关系较近的鲤和金线鲃的分化时间为970万年前(9.7 Mya)。因此,我们推测,鲤特有的异源多倍化事件可能发生于距今970万年至2300万年间。 是否可以将时间更精确一点?试试用转座元件序列吧~ A、B亚基因组的转座元件序列差异(TE divergence)结果显示,鲤两个亚基因组于13%开始分化,在7%时合并形成异源四倍体鲤。由以上已知的A、B亚基因组分化于23 Mya,可将异源四倍体鲤形成时间精确到1240万年前(12.4 Mya),即鲤A、B亚基因组的共同祖先于23 Mya开始分化为两个独立的二倍体物种,之后转座元件伴随着物种分化而发生基因组尺度的序列分化,直到它们伴随着异源四倍化事件(12.4 Mya)重新融合为异源四倍体基因组后,序列分化速率才重新趋于均一。
鲤A亚基因组-鲤B亚基因组、鲤-斑马鱼、鲤-金线鲃同源基因同义替换率 & 亚基因组间转座元件序列差异 | 正文图2d & 2e 亚基因组特征 在多倍体植物中,众多研究显示,其中一个亚基因组倾向拥有更多蛋白编码基因、保留较高的基因表达水平、经历较强的纯化选择(负选择)、维持较低的甲基化水平。这种现象称为亚基因组优势(subgenome dominance)。由此可见,十分有必要对异源四倍体鲤亚基因组基因含量、基因表达模式及表观调控等进行综合分析,以更好理解基因组合并(genome merging)后异源四倍体基因组的演化。 在异源四倍体鲤中是否存在类似植物中的现象? 1 亚基因组基因含量 在比较了鲤两套亚基因组的众多特征后发现,B亚基因组大小和基因含量均显著大于A亚基因组,而GC含量、基因结构、重复元件分布等则没有显著差异。 鲤两套亚基因组的特征参数
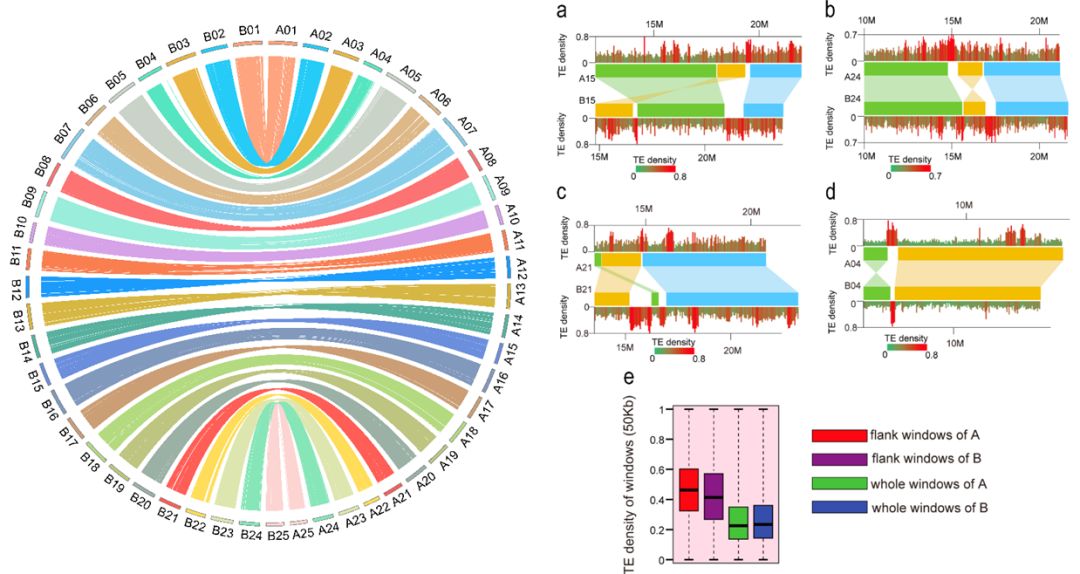
2 亚基因组结构 为了方便下文鲤同源基因命运分化的研究,我们利用可获得基因组数据且最近缘的二倍体草鱼(Ctenopharyngodon idella)作为参考序列,构建了10,724个同源基因对,其中8,291对同源基因(orthologous gene triplets)在草鱼中存在一个拷贝,而在鲤A、B亚基因组分别存在一个拷贝(草鱼 :鲤A :鲤B=1:1:1)。这些同源基因在染色体上的定位极好地展现了两套亚基因组较高的共线性关系(正文图3a)。
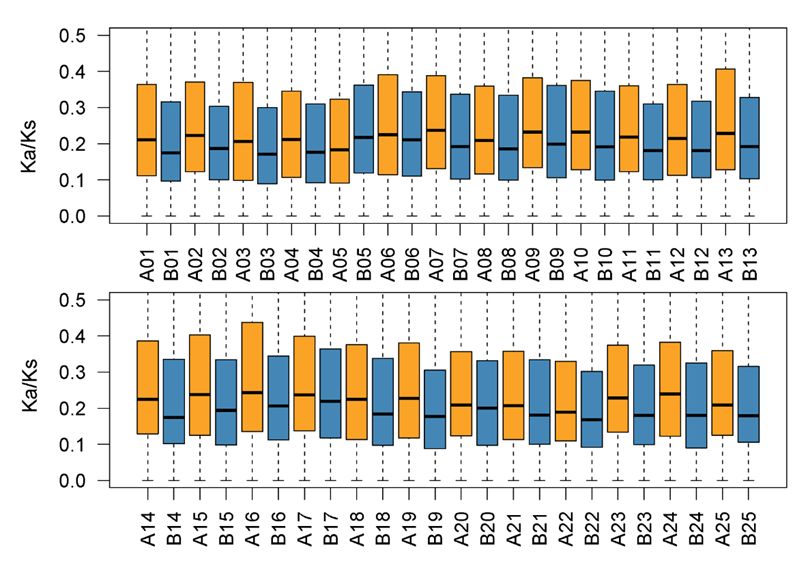
鲤A、B亚基因组共线性、结构变异及对应区域转座元件丰度 | 正文图3a & 附图9 (注:依据近缘二倍体物种基因组测序结果将来自两个祖先的两套亚基因组分开并依据斑马鱼染色体进行重命名,以方便比较研究。) 然而,两个亚基因组仍然存在少量大片段重排(结构变异),包括倒置和易位,如A15和B15染色体存在1.8 Mb的易位、A24和B24染色体存在2.5 Mb的倒置;同时,在结构变异区域周围存在明显的转座元件富集现象(附图9),提示转座元件可能是四倍体鲤同源互换(homoeologous exchanges)的驱动力。 异源四倍体鲤亚基因组不对称演化 以上分析中我们发现,B亚基因组基因含量略高于A亚基因组,这是否足够说明在异源四倍体鲤中,B亚基因组为主效亚基因组?两套亚基因组在选择压力、基因表达模式及表观调控等方面是否有差异? 1 选择压力差异 为了评估两个亚基因组的选择压力,我们计算了同源基因对的非同义突变和同义突变率。明显可以看出,两个亚基因组所受到的选择压力显著不同:B亚基因组经历更强的纯化选择(purifying selection,负选择),A亚基因组则经历较宽松的选择(relaxed purifying selection)。
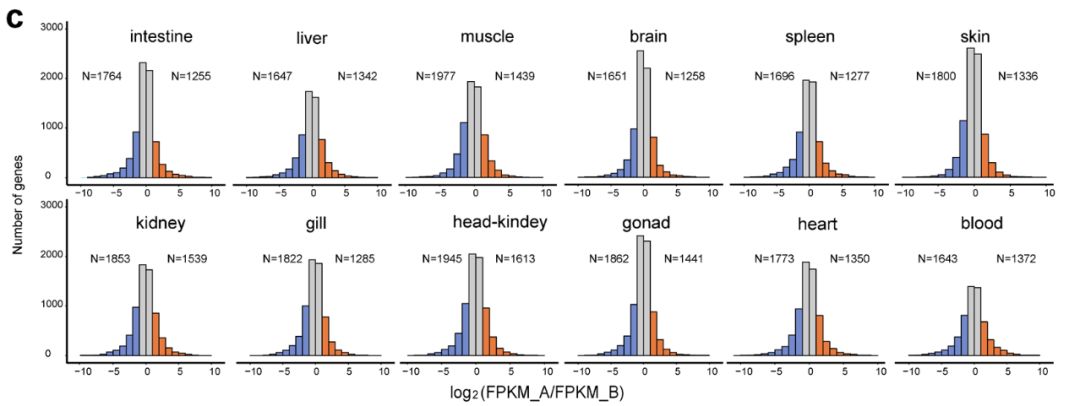
鲤A、B亚基因组共线性、Ka/Ks | 正文图3b 2 基因表达差异 以往在异源四倍体玉米中的研究显示,优势亚基因组较高的纯化选择会使其维持较高的基因表达水平。这种现象在同样是异源四倍体的鲤中是否同样存在?受到较强纯化选择的B亚基因组是否在基因表达水平上具有优势? 带着以上疑问,我们对鲤12个健康组织进行了转录组测序,利用以上获得的8,291对同源基因评估两个亚基因组的表达差异。结果发现,A、B亚基因组存在表达分化,在所有组织中,B亚基因组中表达水平显著高于A亚基因组的同源基因数量均高于A亚基因组中表达水平显著高于B亚基因组的同源基因数量(正文图3c)。这与B亚基因组经历更强的纯化选择一致。 通常情况下,纯化选择倾向剔除有害突变,使基因序列更加保守;而中性选择则会保留适当的变异,使基因功能具有更多可能性,而这种变化通常会降低基因原有表达水平,这也与A亚基因组较松弛的选择压力和较低的基因表达水平一致。由此可见,鲤中B亚基因组在基因表达上是主效亚基因组。
鲤正常组织中A、B亚基因组基因表达差异 | 正文图3c 多倍体生物由于基因组的加倍,使得冗余基因拥有更多向不同功能分化的潜能。在同源四倍体大西洋鲑中,研究显示,多数基因倾向于新功能化,即产生与祖先物种该直系同源基因不同的功能。在异源四倍体鲤中,同源基因又是通过什么样的机制进行演化? 基于以上疑问,我们对近缘二倍体草鱼12个健康组织进行了转录组测序,以其基因表达水平作为祖先物种的参考。对8,291对同源基因进行表达聚类得到3个基因簇:1)鲤A、鲤B均与草鱼直系同源基因存在显著表达差异(306 triplets),提示可能发生亚功能化;2)鲤A与草鱼直系同源基因存在显著表达差异(228 triplets),提示发生新功能化;3)鲤B与草鱼直系同源基因存在显著表达差异(293 triplets),提示发生新功能化。
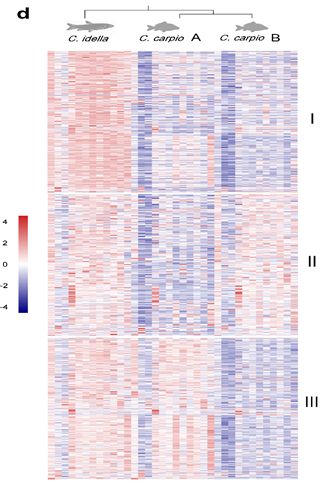
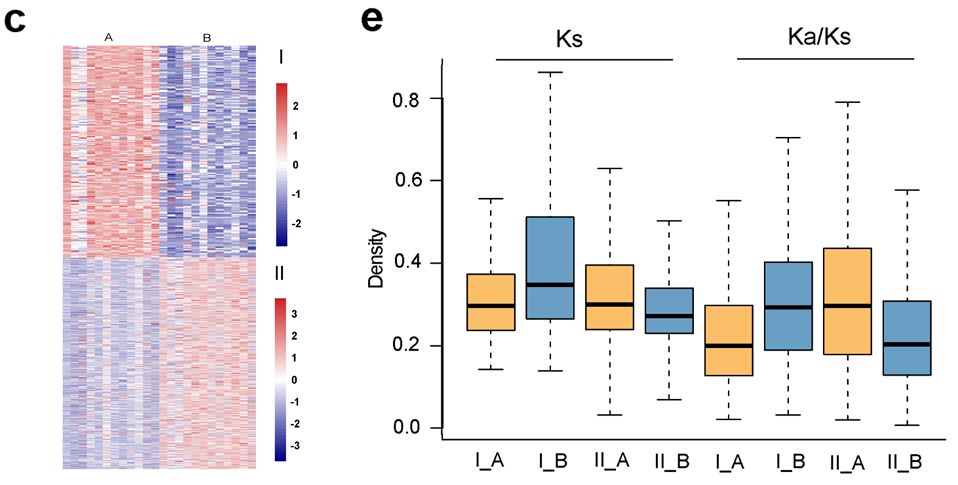
草鱼及鲤A、B亚基因组同源基因表达聚类 | 正文图3d 此外,我们还构建了鲤A、B亚基因组间的表达聚类,发现两个表达模式显著差异的基因簇,其中191对同源基因在A亚基因组高表达,而在B亚基因组几乎不表达;620对同源基因在B亚基因组高表达,而在A亚基因组几乎不表达。这种现象极有可能是同源基因对中其中一个基因发生了非功能化。 什么原因造成了一对同源基因发生如此大的表达分化? 我们对两个基因簇中同源基因进行了选择压力分析发现,无论高表达的基因位于A亚基因组还是B亚基因组,该基因总是具有更强的纯化选择,而表达沉默的同源基因则经历较宽松的选择压力。这也和我们上面提到的,纯化选择通常使得基因功能和表达水平更保守一致。
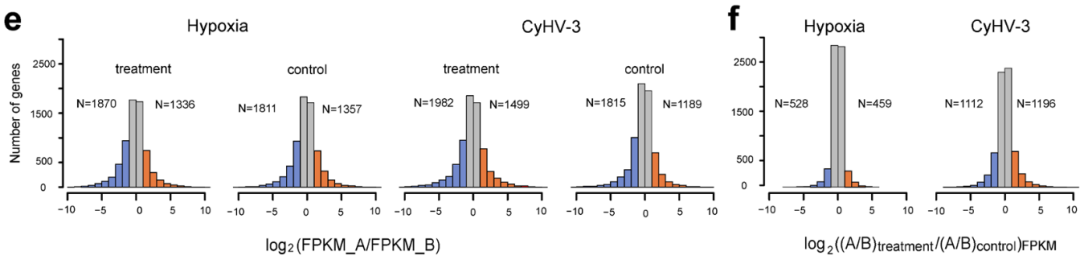
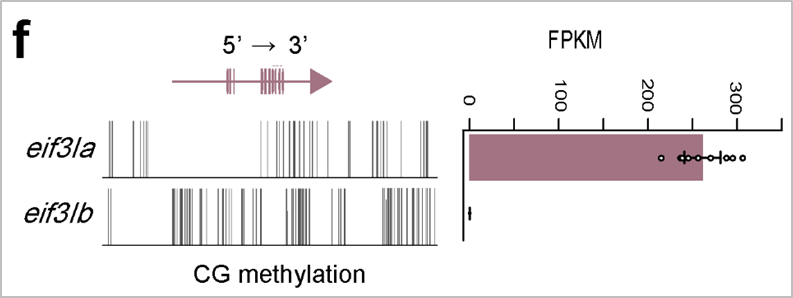
鲤A、B亚基因组表达显著差异的基因簇&不同类别基因甲基化趋势 | 正文图4c & 4e 众所周知,多倍体具有较高的环境适应性。这种在环境适应性上的优势通常是由基因组中多套拷贝基因带来的调控的灵活性。 在鲤中,环境胁迫是否会带来同源基因的表达差异?为了探究这个问题的答案,我们利用公共数据库中获得的鲤低氧(非生物胁迫)、嗜水气单胞菌感染和疱疹病毒感染(生物胁迫)的转录组数据进行一系列分析。结果发现,环境胁迫下两个亚基因组仍然存在表达差异,如eif3I基因位于A亚基因组的拷贝具有高表达,而位于B亚基因组的拷贝则不表达。此外,在整体水平上,环境胁迫会加剧两个亚基因组的表达差异,并且B亚基因组依然具有表达优势(正文图3e & 3f)。
环境胁迫下鲤A、B亚基因组表达差异 | 正文图3e & 3f
eif3I基因的表达和甲基化 | 正文图4f 为了准确反映异源四倍体基因组的环境胁迫响应,十分有必要利用同源基因对的整体表达水平另作分析。我们发现,将一对同源基因作为一个整体分析时,显著响应于该胁迫的基因数量明显降低。由此可见,基因组复制带来了大量冗余基因,增加了异源四倍体基因组的表达可塑性,使其在面对环境胁迫时可通过调控一对同源基因的表达来减少因表达剧烈变化可能带来的不利影响。 本研究可以看出,大多数同源基因仍然保留保守的表达模式和基因功能,因此,在多倍体物种中,利用同源基因总体的表达水平进行转录调控分析,不失为一种新的思路。 3 甲基化水平差异 通常认为,DNA甲基化是一种重要的表观调控机制。而上文我们进行的大量分析显示,B亚基因组为主效亚基因组,具有较高的蛋白编码基因含量、较强的纯化选择压力、较高的基因表达水平。那么,两套亚基因组的这种差异是否和DNA甲基化有关? 由此,我们对鲤进行了全基因组甲基化测序。分析发现,鲤的甲基化位点以CG类型为主,CHG和CHH类型甲基化位点占比极少可以忽略不计。当我们把焦点放在基因编码区域时发现,甲基化改变速率高于碱基替换速率,提示我们,目前阶段的鲤异源四倍体基因组适应性演化可能主要依赖快速有效的表观修饰调控而非不可逆的碱基替换和基因丢失。
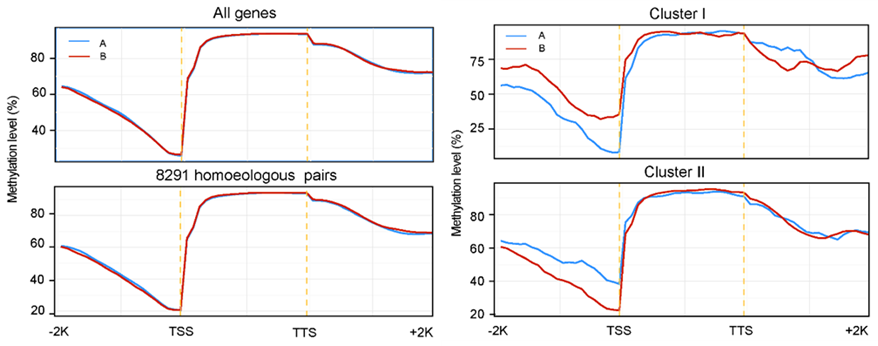
鲤碱基替换与甲基化替换的关系 | 正文图4a 在全基因组水平上,两套亚基因组在基因区和基因下游区域甲基化水平几乎相当,而在基因上游区域可看出主效的B亚基因组甲基化水平略低于A亚基因组。表明基因上游启动子区域的甲基化可能是调控基因表达差异及导致亚基因组优势的原因。
基因组背景下甲基化水平&表达显著差异基因甲基化水平 | 正文图4b & 4d 因此,我们接着研究具有显著基因表达差异的同源基因对,发现在12个组织中,高表达基因上游区域甲基化水平显著低于低表达基因。而对于同源基因表达极端差异的基因,这种趋势更加明显(正文图4c & 4f)。由此可以看出,表观调控对于基因表达分化具有重要影响。 Summary 课题组先后完成了黄河鲤、荷包红鲤和德国镜鲤三个鲤亚种的全基因组序列图谱,并利用鲤高密度SNP分型芯片完成了三个亚种的高密度遗传连锁图谱绘制。 通过对上百种近缘四倍体和二倍体鲤亚科(鲃系)鱼类精细的分子系统学研究,确定了潜在的祖先二倍体类群,随后通过全基因组测序和基因组相似性比较分析,鉴定出“吻孔鲃Poropuntius-小鲃Puntius-裂峡鲃Hampala”为最可能的鲤祖先二倍体类群之一,初步完成了异源四倍体鲤的二倍体祖先溯源,并将鲤异源四倍体起源定位距今1200万年前中新世中期的东亚地区,时间上与青藏高原东部剧烈隆升驱动的气候变化事件高度重叠。 在上述基础上,利用鲤同源染色体序列与近缘二倍体祖先基因组序列的相似性差异,首次成功将异源四倍体鲤基因组拆分为两个各由25条染色体组成的亚基因组,进而评估了鲤亚基因组水平的结构变异、基因和转座元件组成、同源基因(homoeologs)表达分化和表观修饰水平分化。结果表明,鲤异源四倍体基因组仍然保留了完整的两套二倍体亚基因组,并未发现在多倍体基因组中常见的基因丢失和重新二倍化(rediploidization)现象,却观察到大量同源基因不对称表达分化和表观修饰差异,揭示了鲤基因组通过精细调整两套基因的表达水平而非基因突变和丢失来维持正常生命功能的基因组倍性演化模式。 课题组先后完成了黄河鲤、荷包红鲤和德国镜鲤三个鲤亚种的全基因组序列图谱,并利用鲤高密度SNP分型芯片完成了三个亚种的高密度遗传连锁图谱绘制。 通过对上百种近缘四倍体和二倍体鲤亚科(鲃系)鱼类精细的分子系统学研究,确定了潜在的祖先二倍体类群,随后通过全基因组测序和基因组相似性比较分析,鉴定出“吻孔鲃Poropuntius-小鲃Puntius-裂峡鲃Hampala”为最可能的鲤祖先二倍体类群之一,初步完成了异源四倍体鲤的二倍体祖先溯源,并将鲤异源四倍体起源定位距今1200万年前中新世中期的东亚地区,时间上与青藏高原东部剧烈隆升驱动的气候变化事件高度重叠。 在上述基础上,利用鲤同源染色体序列与近缘二倍体祖先基因组序列的相似性差异,首次成功将异源四倍体鲤基因组拆分为两个各由25条染色体组成的亚基因组,进而评估了鲤亚基因组水平的结构变异、基因和转座元件组成、同源基因(homoeologs)表达分化和表观修饰水平分化。结果表明,鲤异源四倍体基因组仍然保留了完整的两套二倍体亚基因组,并未发现在多倍体基因组中常见的基因丢失和重新二倍化(rediploidization)现象,却观察到大量同源基因不对称表达分化和表观修饰差异,揭示了鲤基因组通过精细调整两套基因的表达水平而非基因突变和丢失来维持正常生命功能的基因组倍性演化模式。 迄今为止,二倍体祖先溯源多见于异源多倍体植物,如棉花、小麦、油菜等作物,鲜有在多倍体动物中实现祖先溯源和亚基因组成功分离的报道。这是国际上首次鉴定出异源四倍体鱼类的祖先二倍体并成功进行亚基因组拆分,为开展深入的硬骨鱼类倍性演化和四倍体鱼类性状解析研究扫清了障碍。这些关键科学问题的回答,为深入认识鲤科鱼类多倍体起源演化和探索多倍体鱼类的适应性潜能提供了新的见解,也为在四倍体背景下开展重要经济性状遗传解析,进行基因组选择育种技术创新提供了新的视野。
徐鹏教授与大连海洋大学霍忠明教授进行学术讨论 参考文献: 1. Xu P, Zhang X, Wang X, et al. Genome sequence and genetic diversity of the common carp, Cyprinus carpio[J]. Nature Genetics, 2014, 46(11):1212-1219. 2. 王绪祯, 甘小妮, 李俊兵,等. 鲤亚科多倍体物种独立起源及其与第三纪青藏高原隆升的关系[J]. 中国科学:生命科学, 2016, 46(11):1277. 3. Chen G J, Chang M. A new early cyprinin from Oligocene of South China[J]. Science China Earth Sciences, 2011, 54(4): 481-492. 4. Lien S, Koop B F, Sandve S R, et al. The Atlantic salmon genome provides insights into rediploidization[J]. Nature, 2016, 533(7602): 200. 5. Peng W, Xu J, Zhang Y, et al. An ultra-high density linkage map and QTL mapping for sex and growth-related traits of common carp (Cyprinus carpio)[J]. Scientific reports, 2016, 6: 26693. 内容来源:AGCT与鱼返回搜狐,查看更多 |





【本文地址】