| 下视丘分泌素/食欲素系统之概述 | 您所在的位置:网站首页 › R语言怎么保存数据才能没有行名 › 下视丘分泌素/食欲素系统之概述 |
下视丘分泌素/食欲素系统之概述
|
注:本文来自本人课题组公众号"森林鱼塘",欢迎关注公众号,里面有更多精彩内容!(编者:Can)  1.下视丘分泌素/食欲素的发现 下视丘分泌素(Hypocretin,Hcrt) 在1998年首次被两个不同的研究者同时发现报道。Lecea et al [1] 因发现其主要表达于外侧下丘脑(Lateral Hypothalamus) 且与肠促胰岛素多肽家族(incretin family of peptides)序列相似,故命名为下视丘分泌素。 Sakurai et al [2] 则因发现其具有调节进食的功能而将其命名为食欲素(Orexin),并鉴定了orexin-A 和orexin-B源于同一前体肽,由同一mRNA 编码,对应有OX1R 和OX2R两个受体,OX1R对于orexin-A高亲和,而OX2R不具有选择性。在禁食后,食欲素的表达量显著升高(图1)。  图1 食欲素的发现 图1 食欲素的发现2.食欲素系统的进化保守性 食欲素系统在脊椎动物大脑进化上高度保守,从鱼类、爬行类、鸟类到哺乳类动物中均存在(图2)。  图2 食欲素在进化上高度保守 图2 食欲素在进化上高度保守在所有哺乳动物物种中,食欲素神经元都位于外侧下丘脑(LHA)和邻近区域,包括背内侧和下丘脑后部;在斑马鱼中,食欲素神经元也位于下丘脑的背侧 [3]。 斑马鱼Hcrt神经元网络很简单:在幼鱼中由大约20个神经元组成,在成鱼中大约由60个神经元组成。斑马鱼的Hcrt由一个Hcrt基因编码,它同时编码Hcrt1和Hcrt2肽。斑马鱼上只有一个Hcrt受体,它在结构上更类似于哺乳动物的HCRTR2(表1)。  表1 食欲素系统在斑马鱼,啮齿类动物和人类的对比 [4] 表1 食欲素系统在斑马鱼,啮齿类动物和人类的对比 [4]3.食欲素系统在哺乳动物中的研究 3.1 食欲素系统对睡眠-觉醒状态的调控机制探究 早在1999年,研究者就发现食欲素敲除小鼠表现出嗜睡症,并在其活跃的黑暗期尤其嗜睡,他们的清醒时期是碎片化的 [5]。 2009年有研究发现食欲素能神经元中,GABAB受体的选择性缺失会导致睡眠/清醒结构的中断 [6]。他们将小鼠的食欲素神经元GABAB受体选择性缺失(称为oxGKO小鼠),发现oxGKO小鼠睡眠阶段严重破碎。 随着研究技术手段的进步,研究者使用光遗传学、药理学等方法研究Hcrt神经元和下游去甲肾上腺素能神经元之间的功能连接 [7]。发现在Hcrt刺激过程中,光抑制LC神经元阻断了Hcrt介导的睡眠-觉醒转变(图3)。  图3 LC神经元的抑制阻断了hcrt介导的睡眠-觉醒转变 图3 LC神经元的抑制阻断了hcrt介导的睡眠-觉醒转变研究者在食欲素神经元中选择性敲除5HT1AR,发现在黑暗期的后半段,ox5HT1ARKO小鼠的总清醒时间显著缩短,NREM睡眠时间明显长于对照组小鼠(图4)。该研究阐述了5-HT对食欲素神经元抑制调节的重要作用[8]。  图4 ox5HT1ARKO小鼠在黑暗期后期清醒时间减少 图4 ox5HT1ARKO小鼠在黑暗期后期清醒时间减少总的来说,食欲素具有促进清醒的作用。在促进觉醒的核团中,包括TMN, DR/MnR, LDT/PPT和LC。这些神经元不同程度地表达OX1R、OX2R,并调节清醒、NREM睡眠和NREM-REM睡眠的过渡(图5)。 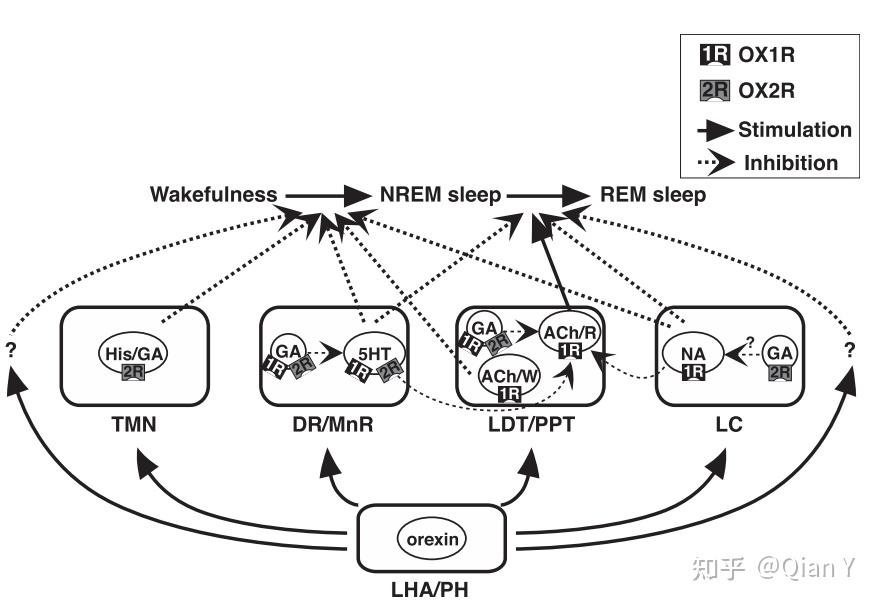 图5食欲素作用的NREM和REM睡眠的示意图 [9] 图5食欲素作用的NREM和REM睡眠的示意图 [9]3.2 食欲素系统对饮食的调控 食欲素的其他功能的研究也有不少报道 [10, 11],CAG/orexin(CAG promoter 过表达)小鼠对饮食诱导的肥胖具有抗性,CAG/orexin转基因减少脂肪量和瘦素,增加能量消耗(图6)。 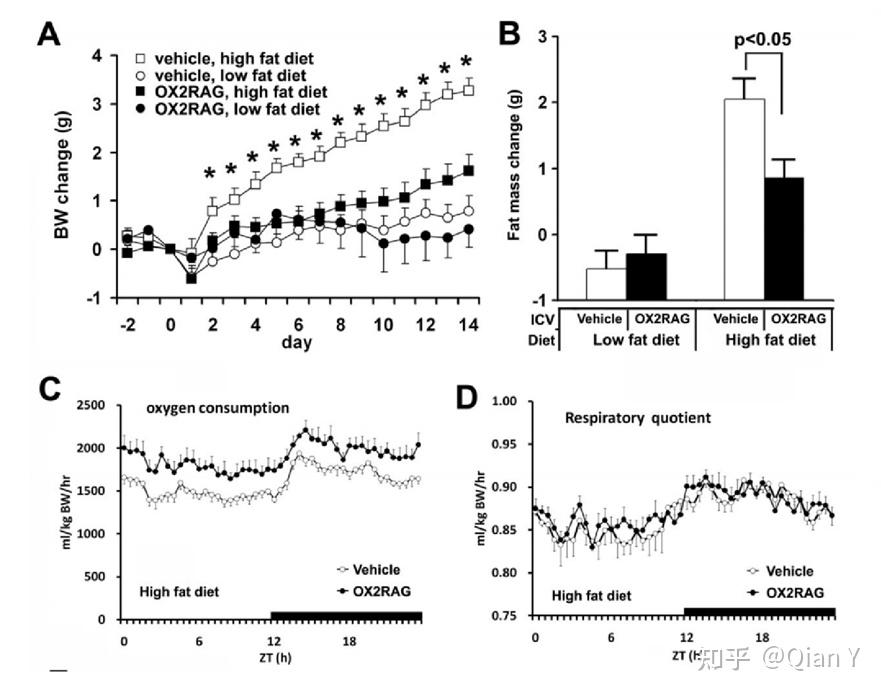 图6 CAG/orexin小鼠对饮食诱导的肥胖具有抗性 图6 CAG/orexin小鼠对饮食诱导的肥胖具有抗性3.3 食欲素系统在学习-记忆中的研究 正常的恐惧条件反射建立需要通过OxR1传递Orexin信号 [12, 13],LC中的OxR1封锁减弱了恐惧记忆的形成,而LC中orexin纤维的光刺激可以增强恐惧记忆的形成(图7)。  图7 正常的威胁学习需要通过OxR1传递Orexin信号 图7 正常的威胁学习需要通过OxR1传递Orexin信号3.4 哺乳动物食欲素的输入和输出神经通路 食欲素神经元的活动受到多重调节,如下图8所示,这些上游输入从行为和环境刺激上来讲,主要包括清醒和皮质唤醒(高伽马脑电图)、探索行为、听觉刺激、情绪刺激、压力和睡眠、食物剥夺。就信号分子而言,主要可分为兴奋和抑制两大类。  图8 食欲素神经元的输入和输出 [14] 图8 食欲素神经元的输入和输出 [14]在2021年发表的食欲素系统(The Orexin System)一书中,对该系统的研究进行了全面系统的总结,已知食欲素和食欲素受体对单胺能系统和胆碱能系统具有调节功能。[15] 4.食欲素系统在斑马鱼中的研究 4.1 斑马鱼食欲素系统及其受体的表达和形态结构的验证 斑马鱼中食欲素的结构和功能相对较少,但也多被证明与哺乳动物在类似。2004年Kaslin等人首次在斑马鱼中鉴定并克隆了Hcrt基因 [16, 17]。原位杂交结果显示,在斑马鱼的大脑中,含有Hcrt mRNA的神经元位于双侧吻侧下丘脑(rostral Hypothalamus),吻侧位于内侧前脑束的内侧,尾侧位于外侧隐窝的顶部 [16](图9)。迄今为止,在斑马鱼中只发现了一种Hcrt受体,这种受体在结构上与哺乳动物的HCRT 2受体相对应 [17, 18]。  图9 胚胎期和幼鱼期Hcrt和Hcrt受体的表达 图9 胚胎期和幼鱼期Hcrt和Hcrt受体的表达斑马鱼Hcrt神经元以谷氨酸能为主 [19]。Hcrt神经元均不表达gad67,少数表达vglut1,多数表达vglut2 (图10)。这也与鼠中的结果是一致的。 在成年斑马鱼中,hcrt纤维分别位于与哺乳动物同源的端脑背侧和pallium下中线。在下丘脑脑室周围区检测到最高密度的轴突投影,HCRT细胞体组织为两个相互相对的紧密簇,没有直接对侧投影 [19]。  图10 斑马鱼HCRT神经元的表达以及成鱼中HCRT系统 图10 斑马鱼HCRT神经元的表达以及成鱼中HCRT系统4.2 斑马鱼食欲素系统对觉醒具有促进作用 Hcrt受体分布于端脑、下丘脑、后结节、松果体和后脑 [17-19]。与哺乳动物中出现的主要的hypocretin能投射到觉醒促进组胺能系统相比,在斑马鱼大脑中的组胺能细胞中还没有发现HCRT受体 [17, 18]。 脑室内注射HCRT1(而非HCRT2)会导致斑马鱼活性的轻微降低,而哺乳动物注射HCRT1后的活性则较强。hcrtR -/-与睡眠有关 [18],而HCRT过表达可以增加斑马鱼在清醒状态下的活动 [17]。 4.3 斑马鱼食欲素核团的输入和输出神经通路 斑马鱼hcrt神经元向大脑发送广泛的上行和下行投射,就像哺乳动物一样。斑马鱼的hcrt神经元投射到单胺能核和胆碱能核 [16-18]。斑马鱼幼体中的Hcrt投影与蓝斑的去甲肾上腺素能细胞和间脑多巴胺能细胞的突起接近 [17]。成年后,Hcrt免疫反应纤维接触这些目标,并密集支配血清素能中脉,可能还支配组胺能和胆碱能类群 [16]。 总结与展望 HCRT是参与许多生理功能的多任务的多肽类,包括调节哺乳动物的摄食行为、清醒和自主-神经内分泌功能以及睡眠-清醒状态。 下丘脑泌素/食欲素神经元的异质性 基于HCRT神经元独特的基因表达谱、连接和活性,研究者认为在所有脊椎动物中,HCRT神经元可分为不同的功能亚群。相对简单的斑马鱼是研究HCRT神经元亚群结构和功能的理想模型。 睡眠-觉醒环路 多个时间尺度上的多个变量的整合是 HCRT功能的必要条件。在HCRT神经元中信息整合是如何实现的?基于层次、冗余和反馈控制的HCRT神经元输入计算模型可以预测睡眠-觉醒的转换,具有较高的精度。食欲素系统之间的联系,它们对清醒和警觉的影响,以及与记忆和认知的联系等仍有待深入研究。 此外食欲素受体也是一个潜在的新治疗靶点,以期改善睡眠障碍、年龄相关性认知衰退和神经退行性疾病的认知等。 参考文献 1.L. DE LECEA, 等人, The hypocretins Hypothalamus-specific peptides with neuroexcitatory activity. PNAS, 1998. 95: p. 322–327. 2.Takeshi Sakurai, 等人, Orexins and Orexin Receptors A Family of Hypothalamic Neuropeptides and G Protein Coupled Receptors that regulate feeding behavior. Cell, 1998. 92: p. 573–585. 3.Soya, S. and T. Sakurai, Evolution of Orexin Neuropeptide System: Structure and Function. Front Neurosci, 2020. 14: p. 691. 4.Sagi, D., L. de Lecea, and L. Appelbaum, Heterogeneity of Hypocretin/Orexin Neurons. Front Neurol Neurosci, 2021. 45: p. 61-74. 5.Richard M. Chemelli, 等人, Narcolepsy in orexin knockout mice molecular genetics of sleep. Cell 1999. 98: p. 437–451. 6.Taizo Matsukia, b., 等人, Selective loss of GABA(B) receptors in orexin-producing neurons results in disrupted sleep-wakefulness architecture. PNAS, 2009. 106. 7.Carter, M.E., 等人, Mechanism for Hypocretin-mediated sleep-to-wake transitions. Proc Natl Acad Sci U S A, 2012. 109(39): p. E2635-44. 8.Saito, Y.C., 等人, Serotonergic Input to Orexin Neurons Plays a Role in Maintaining Wakefulness and REM Sleep Architecture. Frontiers in Neuroscience, 2018. 12. 9.Mieda, M., 等人, Differential roles of orexin receptor-1 and -2 in the regulation of non-REM and REM sleep. J Neurosci, 2011. 31(17): p. 6518-26. 10.Funato, H., 等人, Enhanced orexin receptor-2 signaling prevents diet-induced obesity and improves leptin sensitivity. Cell Metab, 2009. 9(1): p. 64-76. 11.Akihiro Yamanaka, 等人, Hypothalamic Orexin Neurons regulate arousal according to energy balance in mice. Neuron, 2003. 38: p. 701–713. 12.Sears, R.M., 等人, Orexin/hypocretin system modulates amygdala-dependent threat learning through the locus coeruleus. Proc Natl Acad Sci U S A, 2013. 110(50): p. 20260-5. 13.Soya, S., 等人, Orexin modulates behavioral fear expression through the locus coeruleus. Nat Commun, 2017. 8(1): p. 1606. 14.Lawrence, A.J. and L.d. Lecea, Behavioral Neuroscience Of Orexin. Current Topics in Behavioral Neurosciences, 2017. 33(1866-3370). 15.Toor, B., 等人, Sleep, Orexin and Cognition. Front Neurol Neurosci, 2021. 45: p. 38-51. 16.Kaslin, J., 等人, The orexin/hypocretin system in zebrafish is connected to the aminergic and cholinergic systems. J Neurosci, 2004. 24(11): p. 2678-89. 17.Prober, D.A., 等人, Hypocretin/orexin overexpression induces an insomnia-like phenotype in zebrafish. J Neurosci, 2006. 26(51): p. 13400-10. 18.Yokogawa, T., 等人, Characterization of sleep in zebrafish and insomnia in hypocretin receptor mutants. PLoS Biol, 2007. 5(10): p. e277. 19.Lior Appelbauma, b., 等人, Sleep-wake regulation and hypocretin–melatonin interaction in zebrafish. PNAS, 2009. 106. |
【本文地址】