β |
您所在的位置:网站首页 › 酶抑制剂种类和作用特点是什么意思 › β |
β
|
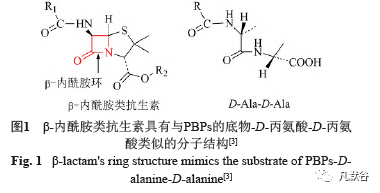
1.2 β-内酰胺药物结构及杀菌原理 关于β-内酰胺药物杀菌原理,国际上有一种公认的理论,认为β-内酰胺抗生素通过抑制参与细胞壁合成的酶,即青霉素结合蛋白(penicillin bindingproteins, PBPs),导致细胞壁受损从而使细菌裂解死亡[1]。β-内酰胺药物可以抑制PBPs,这主要是由β-内酰胺类抗生素的结构特点所导致的。如图1所示,β-内酰胺类抗生素具有共同的环结构,该环与PBPs的底物——D-丙氨酸-D-丙氨酸具有惊人的结构相似性[3]。而早在1970年,Rockefeller University的三位科学家发现,通过抑制一条与PBPs作用途径无关的通路,可以使原本对青霉素敏感的肺炎链球菌对青霉素耐药,这提示我们青霉素杀菌可能存在其他通路。后来发现青霉素让细菌死亡的途径依赖于一种可以降解细胞壁肽聚糖的酶,这种酶就是自溶素[8]。最近有研究对自溶素精确调节肽聚糖降解以及青霉素如何让这一过程失控给出了详细的解答:青霉素通过影响Tacl这种调控自溶素的酶[9],使之表达减少,从而使自溶素从细胞膜转移到细胞壁,最后导致肽聚糖被降解直至细菌死亡[10]。同时,除了抗生素直接杀死细菌的机制外,最新研究发现,不同类型的抗生素在刺激细菌应激时会造成细菌本身活性氧簇(reactiveoxygen species, ROS)持续积累,最终产生或加强杀菌效果。β-内酰胺类药物在作用于细菌时也同样会导致细菌自身ROS不断积累最终达到致死效果,甚至ROS还有帮助氨苄西林裂解细菌的功能[11]。随着科研的不断深入,β-内酰胺类药物导致细菌死亡的途径也不断会有新的发现,可能β-内酰胺类药物在引起细菌死亡时,同时存在着多种途径[3,8-11],但是只要其中一种途径被阻碍时,细菌耐药也可能由此产生。
2 β-内酰胺酶 2.1 β-内酰胺酶的产生 20世纪40年代,对青霉素耐药的金黄色葡萄球菌开始被报道[12]。1940年,人们首次报道了第一种β-内酰胺酶,发现这种酶可以水解β-内酰胺核心,从而使β-内酰胺类的抗生素失效[13]。随着各大制药公司对β-内酰胺药物的加大投入,越来越多的药物进入市场,大大增加了细菌的选择压力,给产β-内酰胺酶细菌提供了独特的生存优势[14],于是人们在多种革兰阴性细菌中发现了β-内酰胺酶,包括大肠埃希菌、肺炎克雷伯菌、铜绿假单胞菌等等。目前已鉴定出超过2000种天然存在的β-内酰胺酶,每种酶都具有独特的氨基酸序列和特征性的水解特性[15]。更糟糕的是,快速复制速率和高突变频率使细菌通过这些β-内酰胺酶的进化以适应新开发的β-内酰胺药物。
2.2 β-内酰胺酶的分类 对于β-内酰胺酶的分类,目前存在两种主要方法,即Ambler和Bush-Jacoby系统[16]。在比较常见的Ambler系统中,首先将β-内酰胺酶根据蛋白质序列和底物特异性分为4类,即A、B、C、D类,再基于其活性位点,将β-内酰胺酶分为丝氨酸-β-内酰胺酶(serine β-lactamases,SBL)和金属-β-内酰胺酶(metallo-β-lactamases,MBL)。SBL包括A、C、D类酶,其需要利用丝氨酸进行β-内酰胺水解。MBL主要是B类酶,其主要需要二价锌离子进行底物水解,但也已报道含有其他二价金属如亚铁离子的MBL具有活性[17]。 根据氨基酸序列、底物特异性和活性位点中金属离子的数量,MBL进一步分为三个亚类:B1,B2和B3[18]。其中B1和B3子类MBL在活性位点均结合两个锌离子,而B2子类MBL在仅结合一个锌离子时最具催化活性,而且当B2子类的活性中心出现第二种金属离子时其活性会受到抑制[19]。最近几年,一种被称为B4子类的全新的MBL在黏质沙雷菌中被发现,其虽与B3型MBL具有同源性但活性位点区域发生了突变:118位氨基酸His变为Arg,121位氨基酸His变为Gln,263位氨基酸His变为Lys。神奇的是,其可以存在单双锌离子两种活性中心状态,既可以以催化惰性的单核酶形式存在,当出现底物时其活性中心被激活,又可以显示双酶活性[20]。 以SBL、MBL系统来说,目前已报道的SBL包括KPC、OXA、SHV、TEM、CMY、CTX、AXC、FOX、TEM和SHV等等,MBL包括NDM、IMP和VIM等等。另外,Bush等[21]于1995年提出基于抗生素药物底物和抑制剂谱对酶进行分组,主要包括第1组(C类)头孢菌素酶;第2组(A类和D类),带有抑制剂抗性,包含超广谱β-内酰胺酶和丝氨酸碳青霉烯酶;第3组金属-β-内酰胺酶。Bush等还建议使用一系列属性来描述新的β-内酰胺酶,包括必需的微生物学特性、底物和抑制剂谱,并且为新的β-内酰胺酶提供有充分代表性的分子序列数据。我们综合β-内酰胺酶的上述两种主要方法及来自Bush和Jacoby团队的数据[16],得出的如表2,我们可以清楚地对应相关酶的活性中心和相关的抗生素底物,这有助于我们更全面地认识这些耐药酶。
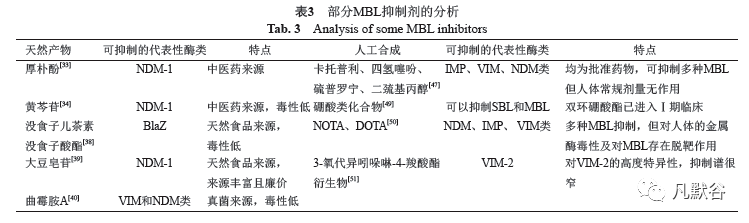
2.3 β-内酰胺酶的检测 新抗生素的开发过程艰难而漫长,对β-内酰胺耐药株进行检测对于控制其感染、拯救患者生命至关重要,同时可以有效减少抗生素的不当使用,从而可以一定程度延长目前可用抗生素的使用时间。目前检测β-内酰胺酶的方法有多种,包括抗菌药物耐药性检测、快速全基因组测序、蛋白组学、电化学发光检测、液相色谱串联质谱多反应检测、环介导等温扩增快速检测法(loop-mediated isothermalamplification ,LAMP)以及新型抗生素的表面结合检测等等方法[22-25]。 传统的抗菌药物耐药性检测需细菌培养,然后进行表型和生化确认测试,实验耗时长。PCR方法目前是检测β-内酰胺酶的金标准[26],当我们从环境或者患者中分离出疑似抗性菌株,将标准的阳性菌株如肺炎克雷伯菌ATCC BAA2473等作为阳性对照,再将结果与之比较。尽管PCR分析属于高灵敏度的分子检测方法,但仍然需要较为昂贵的设备和相关的技术培训,而且PCR技术也相对费时。在2000年,日本东京大学的科学家们开发出了一种称为环介导的等温扩增技术(LAMP),可以在等温条件下高特异性、高效率、快速扩增DNA[27]。这种方法克服了PCR的一些局限性,比如特异性更高,耗时短,无需凝胶电泳即可观察到扩增产物,且因为扩增在等温条件下进行甚至不需要热循环仪[28]。因为在等温条件下(60℃~65℃)进行反应,所以只要在简单的恒温装置内如水浴锅、恒温箱即可进行。其结果的观察也非常简单,只要肉眼看到白色浑浊等产物,即可认为被测物存在靶基因。鉴于以上优点和低成本,LAMP技术适合在经济科技水平较低的国家和地区展开,而这些地区恰恰是因抗生素滥用导致耐药菌产生的热点地区,所以在这些地区推广该检测技术也是十分可行的。 除了基于LAMP这种快速检测方法,最近也报道了一些其他的快速检测方法。由于各国广泛的药物开发研究,最近报道了一种用于快速检测β-内酰胺酶的表面结合抗生素,不同于以往常规需要数天的用于鉴定抗菌药敏感性的方法,该方法能够直接检测尿液中β-内酰胺酶的存在,不过需要利用昂贵复杂的实时分析仪器如PM-IRRAS光谱仪及QCM-D石英晶体微天平进行化学分析[25]。由于诸如液相色谱、质谱等微量检测仪器具有确认蛋白表达的能力,最近也报道了利用液相色谱,串联质谱多反应监测(LC-MS/MS MRM)方法可以在临床分离物中直接检测新德里金属β-内酰胺酶(NDM),且总体测定时间少于90min[22]。 由于几十年来人类不断开发新的β-内酰胺药物,这也导致了对应的β-内酰胺酶的不断出现,当面对新出现的β-内酰胺酶时,我们应如何检测和分型?先进行已知碳青霉烯酶PCR分析发现结果均为阴性,再基于质谱的蛋白组学分析,我们可以将它们识别和分型,如A类β-内酰胺酶Axc[23]。 当然,除了以上提及的检测方法,人们还在开发新的技术以及对现有技术进行联用,达到快速检测的效果。对于β-内酰胺酶的检测,最终还是要根据检测目的以及社会经济与卫生背景因地制宜,选择最合适的方法。 2.4 β-内酰胺酶抑制剂 自从20世纪40年代发现对青霉素耐药的金黄色葡萄球菌开始,到1981年第一次出现阿莫西林-克拉维酸这样的药物抑制剂组合[29],人类探寻抵抗微生物方法的步伐从未停歇。随着新的β-内酰胺酶不断涌现,进入临床的抑制剂也在持续增加,包括克拉维酸与阿莫西林、三唑巴坦与哌拉西林、舒巴坦与氨苄青霉素、头孢他啶/阿维巴坦和美罗培南/瓦巴巴坦等等主要解决了由于广谱β-内酰胺酶ESBLs(集中于A类KPC和某些D类OXA碳青霉烯酶)引起的耐药性[30]。目前,临床上没有对于MBL的抑制剂,SBL的抑制剂对于MBL也无效,我们必须警惕这一状况。MBL几乎对所有β-内酰胺类药物都有抗性,且产MBL的菌株经常呈现出多药抗性表型,并且可以通过质粒(如NDM-1)进行传播[31]。所以我们必须加快MBL抑制剂的研究和开发,争取尽快投入临床,解决日益严重的由MBL导致的耐药感染。目前涉及MBL抑制剂的研究很多,包括人工合成和一些提取的天然产物,它们各有特点,均显示出了一定的开发潜力,有的甚至进入临床试验。 2.4.1 天然产物来源的MBL抑制剂(表3) 就已有的研究而言,可以抑制MBL的天然产物来源很广,包括天然植物、真菌等等,其中天然植物包括传统中医药、食品、植物提取物等是天然抑制剂的主力军。 传统中医药是人类医学瑰宝,从已有的研究中我们也发现了一些具有作为抑制剂潜力的中医药材。厚朴酚提取物(Magnolol)被报道可以抑制NDM-1的活性,在体外实验中得到了证实。厚朴酚是一种从木兰树的树皮中分离出来的天然化合物,与传统的人工合成的化学抑制剂相比,厚朴酚具有来源丰富和易于制备的优点[32]。厚朴酚本身缺乏直接的抗菌活性,而通过杀菌动力学实验发现,其与美罗培南有协同效应,可以恢复美罗培南对于产NDM-1的大肠埃希菌的杀菌活性[33]。通过分子动力学模拟实验,发现厚朴酚可以通过氢键和疏水相互作用与NDM-1结合,从而抑制NDM-1的活性[33]。可以说厚朴酚作为β-内酰胺酶抑制剂的开发,为科学发展传统中医药提供了新思路。黄芩是中华医药宝库中一种有名的药材,有两千多年的用药史。黄芩苷(baicalin)是从其干燥根中提取分离出来的一种黄酮类化合物。其本身具有中等的抗菌活性(MIC≥512μg/mL),当作用于产NDM-1的大肠埃希菌时,黄芩苷将氨苄西林的MIC从128μg/mL降低至8μg/mL,将头孢呋辛钠的MIC从64μg/mL降低至4μg/mL,且由FIC指数所示,与上述两种抗生素有协同作用[34]。同样通过分子动力学模拟实验,发现黄芩苷中的羧基直接与酶活性中心的锌离子相互作用,并且通过其他残基的作用,更稳定地抑制酶的活性。而且,黄芩苷对人的毒性很低,而对NDM-1酶的抑制活性很高。此外,黄芩苷还具有抑制牛乳腺中的大肠埃希菌和降低抗生素耐药性的能力[35],也可抑制铜绿假单胞菌生物膜的形成并增强各种常规抗生素的杀菌作用[36],它也在调节细菌毒力和宿主反应中发挥作用,可作为预防鼠伤寒沙门菌感染的一种很有前景的药物[37]。综合考虑其广谱抗菌性质、与常见抗生素的协同性、对NDM-1酶高亲和力和高抑制能力、对人的低毒性及原料的易获得性,黄芩苷有望作为NDM-1抑制剂药物开发的潜在先导化合物。 在常见食品中也发现一些提取物可以抑制β-内酰胺酶的活性。表没食子儿茶素没食子酸酯(EGCG,儿茶素的主要成分)与青霉素的组合显示出对21种产生青霉素酶的金黄色葡萄球菌临床分离株的协同作用[38],该物质通过直接结合肽聚糖干扰细菌细胞壁的完整性和生物合成而增强了β-内酰胺类药物抗甲氧西林金黄色葡萄球菌(MRSA)的活性,且鉴于绿茶数千年的安全饮用历史,EGCG的毒性很低,从而使体内活性的生物利用浓度的使用成为可能。近几年也发现大豆皂苷对β-内酰胺酶具有抑制作用。大豆皂苷是大豆中存在的两亲化合物,大豆皂苷主要分为A组,以大豆皂甙酚A为糖苷配基;B 组,以大豆皂甙醇B为糖苷配基。浓度低至500μg/mL大豆皂苷与苄青霉素(PCG),奥沙西林(MPIPC)和氨苄西林(ABPC)对MRSA有协同效应,针对不同来源如阴沟肠杆菌、大肠埃希菌的β-内酰胺酶,粗大豆皂苷均可以使包括NDM-1在内的酶活性有效降低[39]。目前虽大豆皂苷的抑制作用的潜在机理尚不清楚,但考虑到其来源丰富、廉价易得,可以说在抑制剂研究和利用方面,都有一定的前景。 在真菌中也发现了MBL的抑制剂。通过亚致死浓度的美罗培南与500多种化合物结合使用作用于大肠埃希菌BW25113ΔbamBΔtolC(大肠埃希菌BW25113通过删除bamB和tolC基因进行修饰,可以提高小分子的通透性并减少其外排)进行筛选,加拿大麦克马斯特大学团队从杂色曲霉菌株(通过18S rRNA基因序列鉴定)的提取物中发现一种真菌天然产物曲霉胺A(AMA)[40],它在体外试验中可以完全恢复了美罗培南对拥有VIM或NDM型等位基因的肠杆菌科细菌,不动杆菌属和假单胞菌属的活性。而且在感染了表达NDM-1的肺炎克雷伯菌的小鼠中,AMA有效地恢复了美罗培南的活性,表明AMA和碳青霉烯类抗生素的组合具有治疗潜力。AMA具有良好的生物耐受性,对小鼠毒性低,对50%受测动物(LD50)致死的剂量为159.8mg/kg,与EDTA相比为28.5mg/kg。所以,AMA很可能是抗生素辅助治疗的极好的潜在药物,有望帮助解决临床上MBL耐药问题。 2.4.2 人工合成的MBL抑制剂 人工合成的MBL抑制剂一直在持续被报道。通常来说,人工合成MBL抑制剂的策略主要有两种:采用锌离子依赖型或非锌离子依赖的抑制剂使MBL失活[41]。锌离子依赖型的抑制剂通过催化酶的活性位点,往往是二价锌离子[42],其主要通过两种方法:①内源性配体的置换以形成可逆的螯合物;②剥离一个或多个二价锌离子使其螯合。 通过配体置换的MBL抑制剂数量十分可观,包括硫醇,硫酮,磺酸,三唑,四唑,异羟肟酸酯和羧酸类化合物[43],其中吡啶二羧酸[44]、吡啶甲酸和吡啶2,4-二羧酸[45]等等也先后被报道。含硫基的卡托普利作为一种常用的抗高血压药,也被报道为MBL的抑制剂,体外试验中其对美罗培南有很好的恢复活性作用,对IMP、VIM、NDM均有良好的抑制作用[46]。通过对结构分析研究,发现其通过游离的硫醇取代了酶的双核桥连锌水分子,为MBL的抑制功能提供了分子证据。除了卡托普利之外已批准的含巯基药物及其衍生物,如四氢噻吩,硫普罗宁和二巯基丙醇,同样具有抑制NDM,VIM和IMP的能力,但可惜的是使用常规剂量这些药物无法恢复实验中亚胺培南的活性,因此主要的优化目标还需提升体内实验中病原体对药物的结合和抗生素活性的恢复[47]。值得一提的是,硼酸类的化合物不仅可以抑制SBL,更显示出了抑制MBL的潜力。因为硼酸盐的四面体结构类似于MBL要水解的β-内酰胺环,可以以水合物形式与活性位点的二价锌离子结合,从而发挥其抑制剂的效果[48]。令人高兴的是,双环硼酸酯(VNRX-5133)与第四代头孢菌素头孢吡肟合用现在正在进行Ⅰ期临床试验[49],期待其早日进入临床并帮助缓解相关的耐药问题。 通过剥离锌离子方法螯合的抑制剂,如已知的金属螯合剂NOTA和DOTA可恢复美罗培南和亚胺培南对表达NDM,IMP和VIM家族成员的细菌的活性,其机制主要是通过充当锌螯合剂,从而使MBL失活[50]。与之原理类似的还有先前提及的来自天然产物的AMA。除了化合物的毒理学限制以外,锌离子依赖的抑制剂还存在着挑战:在人类自身的生化反应中常常存在许多金属依赖性酶,包括锌离子相关的酶,如碳酸酐酶、基质金属蛋白酶和羧肽酶,由于缺乏选择性可能会被这些化合物抑制导致可能的生理不适。除此以外,人类生化循环中如Ca2+和Mg2+相关的酶,在锌依赖性抑制剂作用下这些酶可能与MBL出现竞争抑制从而导致抑制剂的脱靶作用[41]。 基于结构的虚拟筛选方法与N M R ( n u c l e a rmagnetic resonance)过滤相结合,鉴定出VIM-2抑制剂——3-氧代异吲哚啉-4-羧酸酯衍生物[51],这是一种非金属离子依赖型的MBL抑制剂。经晶体学分析,该物质显示出MBL抑制的新模式,即其与活性位点锌离子相邻位点的结合,但是不涉及金属螯合,这样的结果有助于开发新型的临床有效抑制剂。但这种非锌离子依赖型的抑制剂也有着相应的缺点,由于它由VIM-2结构模拟筛选出,因此表现出对VIM-2的高度特异性,对于VIM的其他亚型如VIM-5、VIM-1显示出低抑制活性。这种明显狭窄的选择性可能会限制其用于β-内酰胺酶抑制剂的用途,因为临床上有用的MBL抑制剂应具有针对多种MBL类型(至少是B1亚类)的广谱范围。但是这种抑制剂的优点也相应而生,因为其结合区域不是金属离子中心而是其周围位点,而且MBL核心蛋白折叠存在于人类酶中来抵抗临床上重要的抗癌药物[52],所以这样的抑制剂开发思路对于靶向具有高度选择性的与癌症药物耐药有关的人MBL折叠酶特别有用[53],而且当与适当的测定法结合时,这种选择性抑制剂可用于分析临床观察到的MBL变体[51]。 相比较于已上临床的SBL抑制剂,在本文中我们把更多目光集中在处于探索阶段的MBL抑制剂,我们把上述来自天然产物和人工合成的MBL抑制剂做汇总,得到的表3,希望以这些抑制剂为线索,给抑制剂开发研究带来一些灵感,也给细菌耐药研究带来一些启发。 3 结 语 细菌本身尤其是革兰阴性菌的复杂结构和进化机制,加之人类社会的各种政治经济医疗等等外在条件影响,导致抗生素耐药及超级细菌问题不断被报道。事实上这些问题一定程度上影响着全人类命运,我们必须加以重视,并拿出有效应对措施。随着科技手段和研究的不断深入,对于解决这些问题,新的抗生素靶点甚至新的抗生素开发在进行中,如人们可以通过发现新的抗生素靶点比如最近报道的LiaX[54],从而恢复广泛使用的抗菌剂功效清除多重耐药的超级细菌;一种新型抗生素作用于易位蛋白BamA[55],可对抗多种重要的革兰阴性菌病原体包括大肠埃希菌、肺炎克雷伯菌、铜绿假单胞、鲍曼不动杆菌等等,这些科研成果都为我们对抗超级细菌带来新的曙光。 回到使用历史最悠久的β-内酰胺类抗生素,产MBL的相关菌株可能会更加流行,但通过新的鉴定方法和技术应用,我们会识别出更多的耐药酶,这也为微生物学发展和新药开发提供了丰富的机遇。在多学科交叉的背景下,随着基础研究、政策指导、临床实践的不断深入,我们有理由相信威胁人类健康的β-内酰胺酶引起的耐药问题可以得到有力的控制和解决。
参考文献 详见 中国抗生素杂志 免责声明 我们尊重原创作品。选取的文章已明确注明来源和作者,版权归原作者所有,如涉及侵权或其他问题,请联系我们进行删除。 内容由凡默谷小编查阅文献选取,排版与编辑为原创。如转载,请尊重劳动成果,注明来源于凡默谷公众号。返回搜狐,查看更多 |


【本文地址】
今日新闻 |
点击排行 |
|
推荐新闻 |
图片新闻 |
|
专题文章 |