从个体到集体:基于复杂网络的方法理解蜜蜂的集体行为 |
您所在的位置:网站首页 › 蚂蚁分工合作属于什么行为 › 从个体到集体:基于复杂网络的方法理解蜜蜂的集体行为 |
从个体到集体:基于复杂网络的方法理解蜜蜂的集体行为
原创 Hasenjager等 集智俱乐部  导语 蜜蜂的社会是分工合作的缩影,在这个集体中,个体的利益是如此紧密地结合在一起,以至于在某些情况下,蜂群本身就表现为一个实体。这种非同寻常的自组织需要复杂的通信网络,因此进化出著名的摇摆舞(waggle dance)也就不足为奇了,蜜蜂通过摇摆舞分享关于蜂巢外位置的信息。然而,人们尚不清楚蜂群内的其他信号系统如何协调个体蜜蜂行为,这其中包括颤抖舞(tremble dance)、停止信号(stop signal)和摇晃信号(shaking signal)。这篇文章首先汇集了大量但不相干的历史工作,这些工作调查了这些以上信号对个体蜜蜂的“意义”,然后再讨论基于网络的方法如何分析这些信号,以及这些社交网络如何作为一个复杂的系统来控制蜜蜂社会的集体觅食努力。 关键词:复杂网络,蜜蜂社交网络,集体行为,蜂群 Matthew J. Hasenjager, Victoria R. Franks & Ellouise Leadbeater | 作者 刘志航 | 译者 刘培源 | 审校 邓一雪 | 编辑  论文题目: From dyads to collectives: a review of honeybee signalling 论文链接: https://link.springer.com/article/10.1007/s00265-022-03218-1#Sec7 目录 1. 引言 2. 蜜蜂的信号系统 3. 从个体到网络 4. 讨论 1. 引言 蜜蜂的摇摆舞可以说是动物行为学领域中最著名的通信例子之一,当然也是最迷人的例子之一。早在公元前四世纪,亚里士多德(Haldane 1955)就观察到这种象征性的沟通系统,但直到二十世纪,卡尔·冯·弗里施(von Frisch 1967)才最终破译了这些舞蹈的含义,并为他赢得了1973年诺贝尔生理学奖的一部分。对许多学习动物行为学的学生来说,摇摆舞是一种熟悉的手段,通过这种手段,个体蜜蜂可以减少寻找食物的成本,并与其他个体一起分享一起觅食的好处。在这一研究领域中,鲜为人知的是蜜蜂舞蹈的机制,以及其他交流信号(如颤抖舞:Seeley 1992;停止信号:Nieh 1993;摇晃信号:Schneider and Lewis 2004)和来自花朵气味的嗅觉线索(Farina et al. 2005; 2012)如何控制蜂群的集体觅食行为(Seeley 1995)。摇摆舞不是单独行动,而只是一系列潜在的相互作用的信号网络中的一个信息源,这些信号网络包括正负反馈回路,指导蜂群的行为,从而使集体做出关于如何分配劳动力,以及向何处派遣觅食者的群体“决定”。 虽然这些群体决策的关键信号已经成为近几十年来研究的重点(Seeley 1995;Anderson and Ratnieks 1999;Couvillon 2012;Kietzman and Visscher 2015),但试图研究由此产生的信息交流网络一直具有挑战性。因为追踪一个蜂群中约20,000只蜜蜂中的一小部分,以检测个体信号、线索、反应和典型的行为角色,存在规模上的主要障碍,而分析这种工作中可能产生的海量数据几乎是不可能的(Biesmeijer and Seeley 2005)。但这种情况在近期有所改变,数据处理能力和网络理论的进步都为理解这一复杂系统打开了大门。在这篇综述的前半部分,汇集了关于由觅食的蜜蜂产生的主要运动和振动信号的文献:摇摆舞、颤抖舞、停止信号和摇晃信号。在后半部分,将解释网络理论如何为理解这些信号提供独特的途径,以及现有研究如何将蜜蜂行为从个体推进到蜂群水平。 2. 蜜蜂的信号系统 2.1 摇摆舞 摇摆舞(von Frisch 1967)可以说是蜜蜂中最著名的一种行为(Dyer 2002; Grüter and Farina 2009; Couvillon 2012)。摇摆舞由两个交替的阶段组成:在摇摆运行开始阶段,觅食的蜜蜂沿直线移动,同时剧烈地来回晃动其腹部;返回阶段,进行绕圈,并大致返回到其运行开始的位置(图1a)。每次摇摆运行的持续时间与给定资源以及蜂巢的距离有关,而相对于重力的运行角度与资源相对于太阳的方向有关(von Frisch 1967)。 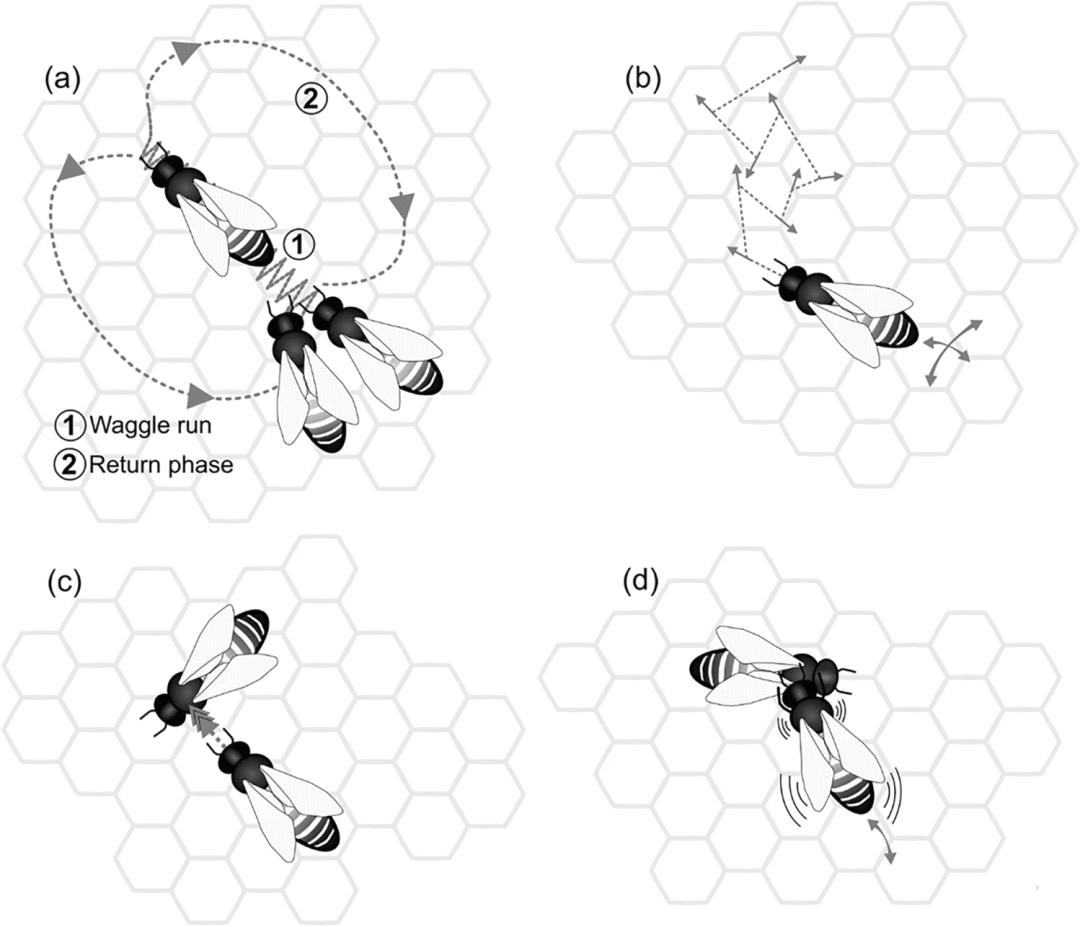 图1. 蜜蜂在集体觅食时使用的四个关键信号。(a)摇摆舞是由信息丰富的摇摆动作组成的,中间有返回阶段(通常是左右交替);(b)在颤抖舞中,蜜蜂以不稳定的轨迹穿过蜂巢,同时振动其身体,有时是前后振动;(c)停止信号包括用头相互撞击,同时发出短暂的声音脉冲;(d)在发出摇晃信号时,工蜂抓住其他蜜蜂,并向背中央摇摆自己的身体。 虽然现在已经确定摇摆舞中编码的空间信息会传递给舞蹈的跟随者(Gould 1975; Riley et al. 2005),但这个假说一度颇有争议。特别是阿德里安·温纳 (Adrian Wenner) 和他的同事们,虽然承认舞蹈的元素与所访问的资源的空间位置相关,但招呼蜜蜂到达食物源有另一种解释:蜜蜂主要通过蜂巢内的嗅觉信息传递来引导食物,它们将这些信息与已知的或寻求的食物点的气味相匹配(Johnson 1967; Wenner et al. 1969; Wenner and Wells 1990; Munz 2005)。最终,冯·弗里施的假说被 James Gould 证实了,在一组实验中,Gould 操纵觅食的蜜蜂,使它们的舞蹈指示出一个它们自己没有去过的地点。足够多的新成员到达所定位的地点,明确地证明蜜蜂能够使用舞蹈传达的信息来定位新的觅食地点(Gould 1975)。 尽管如此,Gould 指出,“只有进一步的工作才能确定在正常情况下,舞蹈语言交流是常见的还是罕见的”(Gould 1975),而且现在已经确定,令人惊讶的是,很少有舞蹈跟随导致到达一个新的食物源(Grüter and Farina 2009)。Biesmeijer and Seeley(2005)通过跟踪觅食蜜蜂一生中的舞蹈跟随,估计在自然条件下,只有 12-25% 的舞蹈跟随互动是为了这个目的。其余 75-88% 的跟随互动被认为是涉及到跟随者已知的食物来源(Biesmeijer and Seeley 2005)。例如,在一段中断的觅食期后(如夜幕降临),跟舞可以重新激活有经验的觅食的蜜蜂,继续从熟悉的位置中采集食物(Johnson 1967; Gil and Farina 2002; Grüter et al. 2008; Hasenjager et al. 2020) 。在这种情况下,舞蹈跟随往往是粗略的(Gil and Farina 2002; Biesmeijer and Seeley 2005; Grüter et al. 2013),表明觅食者不太可能寻求详细的空间信息(Wray et al. 2012)。事实上,蜜蜂经常完全忽略空间信息,跟随一个舞蹈,但随后到达它们自己喜欢的地点(Johnson 1967;Grüter et al. 2008;Hasenjager et al. 2020)。当舞者最终选择寻找新的地点时,它们会进行较长时间的舞蹈跟随(Grüter and Ratnieks 2011;Grüter et al. 2013;Hasenjager et al. 2022),估计是为了获得准确的空间信息。舞蹈跟随,虽然对定位新的觅食区非常重要(Hasenjager et al. 2020),但对于不知道有替代区的觅食者,例如,新手觅食者或当以前访问的地点不再有利可图时(Biesmeijer and Seeley 2005; Grüter and Ratnieks 2011; Grüter et al. 2013),似乎经常作为一个备份信息源。 2.2 颤抖舞 颤抖舞在某些方面类似于摇摆舞,是蜜蜂集体觅食时使用的一种额外信号。当进行颤抖舞时,觅食的蜜蜂迅速沿中轴线摇动身体,并缓慢地穿过蜂巢,频繁地改变方向(Seeley 1992; 图 1b)。颤抖舞被冯·弗里施与摇摆舞一起描述,他认为这个动作没有交流的功能(von Frisch 1967)。然而,虽然颤抖舞被认为没有任何编码信息方面的作用(例如,它们不像摇摆舞那样编码空间信息),但随后从20世纪90年代开始的研究强调,这种信号在交流觅食条件和调节蜂群采蜜处理方面起着关键作用。 Seeley(1992)通过一个经典的实验,为花蜜处理和激发颤抖舞之间的关系提供了重要的证据,他同时训练来自同一蜂巢的两群觅食的蜜蜂到不同的喂食器,然后通过实验增加从一个喂食器采集的觅食者数量。尽管蜂巢外的觅食条件没有发生任何变化,但未经处理的蜂群中的蜜蜂开始颤抖起舞。可能表明这种处理不足的蜂巢内部线索,被认为是发现蜜蜂在到达时卸载花蜜的延迟。事实上,重点蜜蜂队列确实经历了这样的延迟。与此同时,Kirchner 和 Lindauer(1994)发现,以前观察到的一些实验条件也引起了颤抖舞。诸如在食物源处被捕获、遇到拥挤的喂食器或收集到咸的花蜜时,也会导致卸载花蜜的延迟(可能是因为花蜜接收者检测到觅食者身上的报警信息素或不愿意接受味道奇怪的花蜜)。因此,似乎很清楚,卸载花蜜延迟会刺激颤抖舞,Seeley 等人(1996)后来证明,与颤抖舞相关的花蜜流入量增加,随后导致蜂巢工蜂被招呼为花蜜接收者。当一个蜂群的花蜜处理能力因此而提高时,进入的觅食者所经历的延迟应该减少,颤抖舞应该随之停止(Seeley 1995)。 然而,尽管这个反馈回路很优雅,但我们现在知道,蜂巢内部的卸载花蜜延迟不能成为颤抖舞的唯一触发因素。这是因为多达一半的自然颤抖舞在进入蜂巢后立即开始(Biesmeijer 2003b),此时觅食者没有经历任何延迟。许多(但不是全部)舞蹈发生在一天中的第一次觅食旅行中,因此,最近的长期延迟经验不太可能引起颤抖舞的发生。Thom(2003)还表明,在没有卸载延迟的情况下,食物源的拥挤情况可以诱发颤抖舞,这使人们对以下断言产生怀疑(Kirchner and Lindauer 1994):“被捕获、拥挤或其他干扰的蜜蜂(Lindauer 1948)的颤抖舞是它们经历的卸载延迟的一个副作用。”因此,目前还没有提出可以解释所有报告的颤抖舞事件的触发因素,这似乎是对内部和外部觅食条件变化的一种适应性反应,我们只对其功能部分但不完全了解。在本文的后面一节中,我们提出基于网络的方法如何有助于解决这个问题。 颤抖舞也有一个次要的、间接的后果,即会减少摇摆舞的招呼(Seeley 1992),因为一只蜜蜂在觅食后选择颤抖舞,会减少摇摆舞的数量。然而,这种对摇摆舞的负面影响是通过参与调节蜜蜂集体觅食的第三种关键信号——停止信号——更直接地介导的,这种信号与颤抖舞密切相关,下面将介绍。 2.3 停止信号 当发出停止信号时,一只蜜蜂突然用头撞另一只蜜蜂,并通过快速振动其胸部肌肉发出短暂的高调嗡嗡声(图1c)。停止信号的声音成分产生一个独特的波形,其基本频率为350-450赫兹,持续约150毫秒(Kirchner 1993)。至关重要的是,信号接收个体会短暂地“冻结”,这与其他振动信号,如管道信号或乞讨叫声是不同的(Pastor and Seeley 2005)。在觅食的情况下,跳舞的蜜蜂经常也产生停止信号(Nieh 1993),但这种关联并不排斥;事实上,在正常的觅食条件下,大多数停止信号似乎不是由跳舞的蜜蜂产生,而是由最近跟随跳舞的蜜蜂产生(Pastor and Seeley 2005)。它们经常针对摇摆舞者(Nieh 1993; Pastor and Seeley 2005),但产生停止信号的颤抖舞蜜蜂也会将它们指向其他颤抖舞蜜蜂、食物交换者、舞蹈跟随者和其他蜜蜂(Nieh 1993)。 虽然一度被认为是引起食物交换的乞讨信号(Esch 1964; von Frisch 1967),但现在很清楚,停止信号的部分作用是减少觅食蜜蜂的派遣(Pastor and Seeley 2005; Kietzman and Visscher 2015)。首先,发出停止信号的蜜蜂很少从它们的目标蜂群伙伴那里得到食物(Nieh 1993)。第二,当停止信号针对摇摆舞者时,它们会停止跳舞并离开(Kirchner 1993; Nieh 1993);这种反应也可以由与停止信号相同频率产生的合成声音触发(Nieh 1993)。第三,也有一些来自一个蜜蜂物种(A. cerana)的证据表明,守卫蜜蜂在巢穴入口处产生的停止信号可以抑制工蜂的离开(Tan et al. 2016)。 颤抖舞的蜜蜂经常发出停止信号(Nieh 1993),这与作为紧急刹车器的作用是一致的,它打断了由摇摆舞招呼产生的正反馈循环,使蜂群有时间在招呼恢复之前建立花蜜处理能力。同样,当觅食者在食物竞争期间遇到同种攻击、捕食者攻击或存在报警信息素时,停止信号的产生急剧增加(Nieh 2010;Jack-McCollough and Nieh 2015;Tan et al. 2016),并在危险更大时变得更强烈,这也是有道理的(Tan et al. 2016;Dong et al. 2019)。例如,在抢夺花蜜的事件中,如果目标蜂群被证明有能力保护自己,那么停止抢夺蜂群的招呼可能对限制攻击相关的成本至关重要(Johnson and Nieh 2010)。然而奇怪的是,这种情况下通常也会引起颤抖舞的产生。如果颤抖舞是为了招呼花蜜处理能力,为什么它发生在引起停止信号的捕食/竞争背景下?如果针对其他颤抖舞者、花蜜接受者、舞蹈追随者或其他非觅食蜜蜂的停止信号具有某种功能(Nieh 1993),那么它是什么?解决这些问题可能具有挑战性,因为它们需要很好地评估信号发出者和接受者在信号互动之前所从事的任务,并持续跟踪以确定随后的行为变化。对个体的自动跟踪,结合可以分配行为角色网络的分析,现在提供了一种应对这一挑战的手段,我们将在下面进一步讨论,在我们介绍了有助于觅食的最后一个信号:摇晃信号。 2.4 摇晃信号 与上述信号不同的是,摇晃信号通常会引起接收者行为的相对直接和具体的变化,摇晃信号被认为是一种调节性的交流信号,它可以改变接收者的整体兴奋性或活动,但不会导致其行为的一个具体变化(Nieh 1998;Schneider and Lewis 2004)。摇晃信号是由信号发出者用它的前胸和中胸腿抓住另一只蜜蜂,然后以大约16赫兹的频率继续振动自己的身体背中央1-2秒(图1d)。然而,确定摇晃信号所传递的信息已被证明具有挑战性,因为它是在几种情况下产生的(例如,觅食的开始和停止、蜂群、蜂后饲养),它对接收者的影响似乎是高度特定的,取决于接收者的生理学、年龄、任务表现和遗传学(Schneider and Lewis 2004; Koenig et al.2020)。例如,摇晃信号的接受者移动得更快,并显示出细胞活率的提高、视线移动、摇摆舞动和梳理行为(Nieh 1998;Cao et al. 2007;Koenig et al. 2020)。此外,一些行为效果在收到信号后15-30分钟才明显(Schneider and Lewis 2004)。然而,有几条证据表明,摇晃信号在调节集体觅食活动中起着重要作用。 在觅食方面,大多数摇晃信号是由觅食者产生的,往往是在觅食成功之后(Nieh 1998; Biesmeijer 2003a)。摇晃信号通常是在一天中第一次成功觅食后产生的,这时蜂巢中的许多蜜蜂仍然相对不活跃(Seeley et al. 1998)。此外,前几天经历过觅食成功的蜜蜂,甚至在开始觅食前就会产生摇晃信号(Biesmeijer 2003a)。因此,摇晃信号的产生有可预测的昼夜变化,信号活动的高峰在摇摆舞的产生之前(Nieh 1998; Biesmeijer 2003a; Ramsey et al. 2018)。在蜜蜂经历了食物匮乏后,摇晃信号的产生往往特别明显(Seeley et al. 1998)。这些觅食者产生的摇晃信号的接收者往往是年轻和/或不活跃的蜜蜂(Lewis et al. 2002;Nieh 1998),然而如上所述,接收者的反应可能是高度异质性的。 摇晃信号的一个可能功能是,通过增加接收者的运动和活动,摇晃信号增加了与其他蜜蜂和蜂群内其他任务相关信息来源互动的可能性(Biesmeijer 2003a;Cao et al. 2007)。因此,摇晃信号的接收者将更快地获得关于蜂群状态和食物可用性的最新信息。因此,摇晃信号可以提供一种间接手段,觅食者可以通过这种手段促进工作的重新分配,以应对觅食条件的变化。事实上,尽管通常在清晨开始觅食时观察到摇晃信号的高峰,但也可以观察到下午的小高峰(Nieh 1998)。这些下午的摇晃信号可能有助于协调群落觅食和食物加工活动的停止,促进接受者重新评估当前的活动(可能通过增加活动来介导;Nieh 1998)。然而,与颤抖舞和停止信号一样,要更好地了解摇晃信号在协调群落觅食方面的作用,还需要有能力在较长的时间内跟踪信号发出者和接受者的身份、工作历史和社会经历。我们现在要讨论的是,为研究复杂的网络而开发的工具,加上追踪技术的进步,如何为应对这些挑战提供潜在的有用手段。 3. 从个体到网络 虽然上述信号,在大量的嗅觉线索中(von Frisch 1967;Farina et al. 2005;Thom et al. 2007),允许个体蜜蜂更快地找到食物,招呼花蜜接收者,并在需要时停止觅食,但它们也涌现出了蜂群层面的觅食行为,超出了个体信号配对的总和(Sumpter 2006)。为了提供一个有点抽象的集体决策概念的例子,考虑一下蜂群在花蜜源之间的选择,而不需要任何一个个体去单独比较觅食地点。当一只蜜蜂找到超过阈值浓度的花蜜时(阈值由季节性食物可用性决定;von Frisch 1967),它在返回蜂巢时的舞蹈表明了它的位置,但也反映了这次觅食之旅的能量效率。这是因为更有效率的旅行(效率来自旅行和收集信息时间,由收集的花蜜的热量回报来计算)比效率低的旅行引起更多的舞蹈(Seeley 1994)。因此,舞蹈跟随者更有可能遇到这些食物源的舞蹈,因此更有可能被招呼到这些食物来源,而不需要任何个体判断某个特定的食物来源比其他食物来源更好(Seeley and Towne 1992)。蜂群集体,而不是个体,决定在哪里分配劳动力,集体倾向于更近和更甜的蜜源。 一个类似的例子可以在蜂群选择巢穴的情况下找到,摇摆舞者不仅为它们找到的潜在巢穴跳舞(为更高质量的巢穴表演更多的舞蹈,就像觅食一样;Seeley and Visscher 2008),而且还对那些宣传替代方案的舞者发出停止信号(Seeley et al. 2012)。结合起来,这两个信号网络实现了一个结果,即信息在群体中迅速流动,允许迅速停止对劣质场地的招呼,并减少在类似场地上出现僵局的可能性。虽然建立这种群体层面行为的基本单位是个体的配对。例如,一个摇摆舞者和一个后来可能成为舞者的舞蹈跟随者,但群体本身是一个复杂的系统,通过这种信号产生的网络的特性而建立。 近年来,网络理论在动物行为中的应用极大地促进了人们对复杂社会系统的结构及其对行为、生态和进化过程的影响的理解(Hasenjager and Dugatkin 2015;Krause et al. 2015)。网络理论提供了一个框架,用于描述相互作用的主体系统(如工蜂),作为由表示某种形式关系的发生和强度的边连接起来的节点集。就社会性昆虫而言,网络的例子包括信号提供者和接受者之间的互动(例如蚂蚁的出行:Richardson et al. 2018年)、触角接触(Gordon 2010),甚至协同互动(通过环境改变进行间接交流;Richardson and Gorochowski 2015)。以这种方式表示社会系统能够使用数学工具来确定其关键要素是什么,以及它们的结构如何塑造整个系统的行为(Waters and Fewell 2012;Pinter-Wollman et al. 2014;Farine and Whitehead 2015)。虽然网络方法的一个突出缺点是对数据的要求非常高(Farine 2018;Finn et al. 2019),但最近自动跟踪动物行为的技术的进步意味着,现在可以获得关于整个蜂群中个体行为的全面的高分辨率数据(Buenisch et al. 2018;Gernat et al. 2018;Wild et al. 2021),包括自动检测特定的交流互动(如交哺:Gernat et al. 2018;Wild et al. 2021;摇摆舞:Wario et al. 2017;Okubo et al. 2019;摇晃信号:Ramsey et al. 2018)。 在基于网络的工具箱出现之前,当然可以探索社会性昆虫的集体行为(Heinrich 1981;Deneubourg et al. 1990;Seeley and Buhrman 1999;Mallon et al. 2001)。因此,我们有理由问,网络科学是否能有助于理解蜜蜂信号网络的进化功能,而不是简单地描述产生蜂群行为的近似机制。我们认为它可以,原因如下:首先,正如前面几节所强调的,目前的经验方法在描述一些信号的基本生物“意义”方面已经达到了极限(例如,接收者对颤抖舞和摇晃信号的反应往往是高度异质性的;Biesmeijer 2003b;Schneider and Lewis 2004)。网络方法可以使详细的互动历史有意义,以补救这个问题。与此相关的是,解释为什么系统中存在这么多明显的冗余,需要了解信号类型是如何相互协同、叠加或对抗的,而网络工具已经被开发出来,可以分解这些影响(De Domenico et al. 2016;Hasenjager et al. 2021a)。最后,网络本身是群体层面的行为表征,选择可以塑造它们以实现特定的目标。网络科学提供了各种指标来捕捉这些结构特征(见表1;Farine and Whitehead 2015),可以说明选择是如何促进(或减少)群体内的信息流、疾病传播或劳动分工等过程的,这在个体水平的方法中是不明显的(Otterstatter and Thomson 2007;Pinter-Wollman et al. 2011;Mersch et al. 2013;Hasenjager et al. 2020;Easter et al. 2022)。我们在下文中逐一展开,强调如何使用目前的网络方法,介绍已经开始探索这些方法的研究,并讨论为蜜蜂系统带来更适合的方法的扩展。  3.1 将信号与结果联系起来 推断一个信号的功能,首先需要了解谁产生了这个信号,它是针对谁的,以及它如何改变他们的行为。当互动涉及以蜂巢为基础的蜜蜂时,这可能是具有挑战性的,因为它需要了解互动前的行为历史,因此,年龄通常被用作行为角色的属性,基于工蜂内部的时间多态性(Free 1965)。然而,这种方法不能考虑到工蜂之间在发育轨迹上的巨大差异,这种差异反映了遗传学、育雏要求和外部条件的不同。为了弥补这一点,Wild 等人(2021)使用自动跟踪来跟踪一个蜜蜂群25天,重建个体的日常互动网络,并利用它们来确定每只蜜蜂在不同时间点的精细行为角色。重要的是,仅仅标记蜂群的 1% 就足以在推断个体的行为角色方面取得强大的性能,而在 5% 时,性能相当于标记整个蜂群所取得的性能。Wild 等人的方法提供了一种手段来识别暴露于特定信号后活动模式的变化,如重复的颤抖舞、停止信号或摇晃信号。例如,这种方法可以被证明有助于探索那些传递给非觅食蜜蜂,或由其产生的停止信号的功能,或那些在觅食情况下产生的颤抖舞。这种方法还可以促进对摇晃信号如何影响任务分配的分析,因为它们对接收者行为的影响被认为在很大程度上取决于个体过去的经验,并且可能在互动发生后一段时间才表现出来(Schneider and Lewis 2004)。然而,Wild 等人的方法最令人兴奋的潜在应用在于它有可能捕捉信号之间的相互作用,即一个信号的影响取决于蜜蜂是否收到另一个信号。通过提供每个个体的整个行为历史,可以研究信号之间的协同和对抗作用(例如使用多层网络框架,我们将在下一节讨论)。 网络科学还提供了一种量化对信号反应的手段,通过一种叫做基于网络的扩散分析(NBDA;Franz and Nunn 2009;Hoppitt et al. 2010)的技术。NBDA 的核心假设是,如果一个特定类型的互动能够传递信息,那么相应的互动网络应该预测个体开始表达相关行为特征的顺序。例如,在其他条件相同的情况下,对于一个给定的地点,跟随更多摇摆舞线路的觅食者(因此在跟随舞蹈的网络中联系更紧密)预计会比跟随较少线路的个体更早找到该地点。到达的模式越是紧跟这个舞蹈跟随网络,估计舞蹈跟随的社会传播效应就越强。重要的是,多种网络类型可以包括在 NBDA 中,使研究人员能够解决它们对特定反应的相对贡献(表1)。例如,尽管早已确定蜜蜂能够利用摇摆舞中的空间信息,但同样确定的是,嗅觉线索也是引导蜜蜂走向食物的关键(Gould 1975;Johnson 1967;Riley et al. 2005;Grüter et al. 2008),我们最近采用 NBDA 来量化摇摆舞交流和嗅觉线索在帮助工蜂找到新的觅食地点方面的相对贡献(Hasenjager et al. 2020)。 为了区分基于舞蹈和嗅觉交流途径的影响,我们对一组觅食者的巢内互动进行了录像,这些觅食者熟悉的喂食器已不再提供奖励,而另一个新的喂食器正在蜂巢中进行宣传。从视频记录中,我们构建了捕捉摇摆舞互动的网络和基于传递嗅觉信息的互动的独立网络(如交哺和触角感触)。仅仅摇摆舞的相互作用就能够解释 97% 的新喂食者的招呼事件,突出了尽管同时有嗅觉线索,但舞蹈在发现食物来源方面的关键作用(Hasenjager et al. 2020)。相反,当我们在重新发现一个熟悉的喂食器的情况下比较相同的网络时,嗅觉线索获得了更大的重要性,在解释喂食者的到达方面变得与舞蹈同样有效(Hasenjager et al. 2020)。 传统上,网络分析往往依赖于静态网络,忽略了互动发生的顺序——这种方法在研究例如稳定的社会关系时有意义,但在研究信号互动的后果时却没有意义(Blonder et al. 2012;Farine 2018)。因此,将 NBDA 应用于蜜蜂信号,需要我们使用可以随时间变化的网络(Hasenjager et al. 2020;2021a)。已经开发了两种主要的方法来模拟时间网络:时间聚合网络,基本上是一系列的静态网络,和时间序列网络,产生所有时间点的所有相互作用的完整记录(Blonder et al. 2012;Hasenjager et al. 2021a)。哪种方法更有用,往往取决于研究问题。例如,时间聚合网络可用于评估特定的蜜蜂觅食者是否在多日的信号(如舞蹈——跟随)网络中始终占据中心位置,而调查接收者在调节觅食者信号的生产持续时间或功效方面的潜在作用,可能会受益于更细化的时时间序列方法(表1)。也可以将时间分类网络表示为多层网络,其中每一层代表不同的时间窗口(Finn et al. 2019;Fisher et al. Pinter-Wollman,2021)。我们在下一节讨论捕捉不同信号组合的多层网络的概念。 3.2 了解互动效应 对理解信号“意义”的传统方法来说,一个问题是,一个信号的影响可能取决于个体已经收到的其他信号。例如,Thom 等人(2003)假设,停止信号有时会起到降低接收者对颤抖舞的反应阈值的作用。同样,个体跟随摇摆舞的可能性可能取决于它以前是否收到过摇晃信号(Nieh 1998),而摇晃信号又往往是由产生摇摆舞的相同个体产生的。记录这些类型的协同、对抗或相加的互动作用,对于理解高度社会化群体内的交流如何演变是至关重要的。正如我们上面所描述的,自动跟踪所带来的详细的互动历史(Wild et al. 2021年)将是这里的关键,但能够理解这一庞大信息的工具也是至关重要的。 为此,动物网络研究不再局限于所有连接代表相同形式的相互作用(如交哺,空间邻近)的网络,或者从多种相互作用类型中产生一个聚合的行为测量。相反,多层网络方法将多个网络结合成一个数学结构,同时保持每个网络类型(即层;Silk et al. 2018;Finn et al. 2019;Hasenjager et al. 2021a)的独特性。例如,可以构建一个多层网络,其中每一层都是捕捉不同互动类型(如摇摆舞、颤抖舞、停止信号)的网络,各层的个体与自己相连(图2)。多层网络框架使研究人员能够解决使用传统单层方法无法获得的问题(表1)。例如,人们可以问:蜜蜂个体在通过停止信号对基于摇摆舞的招呼施加负反馈方面的作用,是由其在停止信号层的网络位置单独捕捉到的,还是由同时包含其在停止信号层的连接和其目标在摇摆舞层的连接信息的多层中心性测量(如特征向量多功能性;De Domenico et al. 2015)捕捉到的?换句话说,只有停止信号者需要有良好的连接性才能有效地传递其信息,还是他们也需要其摇摆舞的接受者有良好的连接性?多层网络方法也可以用来研究各层之间的互动动力学耦合有多强,以及这种耦合的强度是否因环境而异(Bianconi 2013;表1)。例如,颤抖舞和停止信号都与减缓觅食者的招呼有关(Seeley 1992;Nieh 1993),但似乎也有其他潜在的不相关功能(Nieh 1993;Seeley et al. 1996)。比较这些网络在不同背景下的结构,可以突出它们功能一致或不同的条件。最后,不同网络类型的多层表示可以进一步与捕捉每个时间窗口的一组额外的层结合起来(图2),从而能够分析在一个时间点上的相互作用(如颤抖舞,停止信号)如何串联起来影响随后时间点上层内和层间的互动作用模式(如摇摆舞——跟随)(Bonnell et al. 2021;表1)。 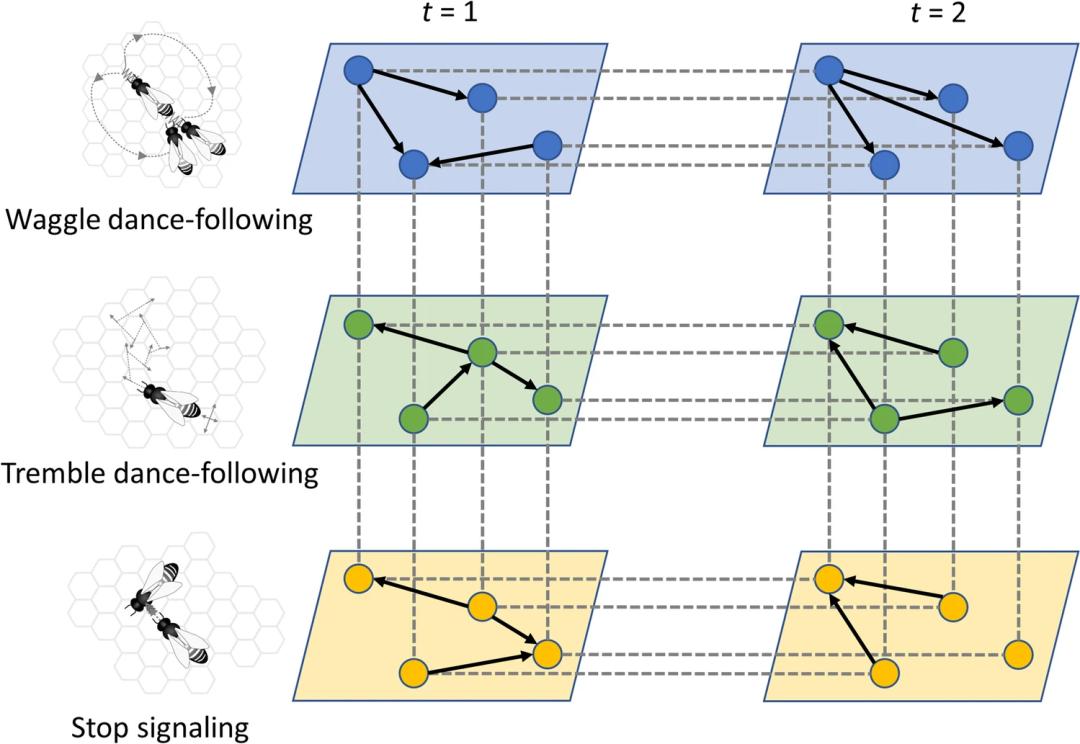 图2. 一个由四个个体组成的多层网络,包括三个行为和两个时间段。在一个特定的层中(用阴影的平行四边形表示),个体用节点表示,它们之间的相互作用用实线表示。每一行对应于不同的互动形式(摇摆舞、颤抖舞、停止信号),每一列对应于时间段。个体通过层间边(虚线)在时间段内和跨时间段与自己相连。 3.3 蜜蜂网络的适应性结构 正如我们所看到的,蜜蜂觅食者所使用的信号并不只是在配对者之间传递信息,而是使群体层面的行为得到协调,例如将觅食者的采蜜率与蜂群处理传入花蜜的能力相匹配(Seeley et al. 1996)。选择可能作用于集体活动的一个更微妙的方法是通过互动网络本身的特性,它产生于工蜂之间互动的模式化(Gordon 2010; Naug 2015)。例如,特定年龄段的蜜蜂之间的差异化互动可以导致网络中出现子集群,从而决定传播动力学,并可能使信息(例如关于觅食地点的信息)针对觅食年龄段的群组,同时隔离年轻的蜜蜂,这些信息对它们来说不太相关,它们可能暴露于通过这些相同的互动途径传播的病原体(Naug 2008)。 个体之间的行为变化也经常与增强的集体结果相关,包括蜜蜂和其他社会性昆虫的觅食率和蜂群生产力的提高(Jandt et al. 2014;O'Shea-Wheller et al. 2021)。网络理论提供了各种指标,可以捕捉这种变化并将其与群体层面的行为联系起来(Farine and Whitehead 2015;表1)。例如,特征向量中心性既说明了个体的直接联系,也说明了其网络邻居的联系程度,因此可以反映一个蜜蜂在整个网络中传递信息的重要性。这类指标可以进一步与模拟模型相结合,预测和评估观察到的变异模式如何影响蜂群动力学的潜在机制(Gordon 2010;Gernat et al. 2018)。例如,以前的研究表明,高度互动的个体可以作为网络枢纽出现,对信号传播有加速作用(Pinter-Wollman et al. 2011)。如果这种“枢纽”式的个体存在于不同的信号网络中,具有潜在的协同效应(如颤抖舞和摇晃信号)或对抗效应(如摇摆舞和停止信号),对集体进程可能会产生什么后果(表1)?从模型中产生的预测随后可以通过操纵工蜂变异和/或信号产生的实验来检验(例如,去除高中心性与低中心性的觅食者,Crall et al. 2018;人工生产信号,Koenig et al. 2020)。多层网络方法还能进一步比较信号系统内部和之间的个体变化。由不同信号系统构建的网络是否具有相似的组织性?一个觅食者在网络中的重要性是相关的还是在不同信号层中高度可变的?一个蜜蜂在网络中的中心地位在一段时间内有多稳定? 识别群体相互作用网络结构中的功能元素的最后一种有希望的方法,是寻找跨个体子集的重复出现的模式或模体(Milo et al. 2004;Easter et al. 2022)。一个这样的模体是前馈环路,其特征是两个节点(如A和C)之间的一对定向路径,涉及直接路径和由第三个节点介导的间接路径(即A到C和A到B到C的分别)。前馈环路可能非常适合通过网络有效地引导信息流来调节群体行为(Mangan and Alon 2003)。事实上,在收割蚁 Pogonomyrmex californicus 的触角网络和黄蜂 Ropalidia marginata 的优势网络中都发现了这种模体,可能反映了这些网络在调节蜂群功能方面的作用(Waters and Fewell 2012;Nandi et al. 2014)。同样,通过在蜜蜂觅食者的信号网络中寻找前馈环路和其他模体,可能会深入了解网络的功能特性以及选择如何作用于觅食者行为。 4. 讨论 在进化层面上,蜜蜂集体觅食的最终价值仍然是一个值得争论的话题(I'Anson Price et al. 2019)。事实证明,很难确定摇摆舞——作为集体决策基础的关键通信系统——能够提高蜂群的觅食成功率。这样做的尝试通常涉及到使舞蹈提供的空间信息变得毫无意义的操作,这有时会对更多零星的、具有挑战性的景观中的食物采集产生负面影响(Sherman and Visscher 2002;Dornhaus and Chittka 2004;Donaldson-Matasci et al. 2013),但情况并非普遍如此(Donaldson-Matasci and Dornhaus 2012;Nürnberger et al. 2017;I'Anson Price et al. 2019)。这样的操作在规模上是费力的(Nürnberger et al. 2017),而确定那些舞蹈追随者选择使用舞蹈中的信息的情况,可能代表了一个富有成效的替代方案。例如,正如我们上面所描述的,Hasenjager 等人(2020年)使用 NBDA 表明,当蜜蜂在相似的食物来源之间移动时,舞蹈会驱使它们去寻找食物;将这种范例扩展到气味不同的食物来源之间的过渡(模仿物种之间的转变),可能会发现舞蹈不那么重要,因此突出了物种内(比方说物种之间)不同开花树木之间快速转变的需要,这是舞蹈进化的潜在动力。 虽然目前用于长期跟踪蜜蜂行为的计算机视觉方法为了解蜜蜂的社会生活提供了前所未有的机会(Wario et al. 2017;Gernat et al. 2018;Wild et al. 2021),蜜蜂使用的许多信号和线索不能被基于视觉的方法捕获(Seeley 1998),包括觅食者产生的信息素(Thom et al. 2007),蜂王广播的声音信号(Michelsen et al. 1986)和飞行或其他活动中积累的静电荷(Greggers et al. 2013)。开发能够将这些和其他形式的互动整合到网络模型中的方法,可以进一步提高对蜜蜂集体组织的理解。潜在的有前途的方法包括将视觉跟踪与其他感官模式(振动:Ramsey et al. 2018;静电:Paffhausen et al. 2021)的自动监测结合起来,纳入协同互动(Richardson and Gorochowski,2015),以及基于网络方法的扩展,明确允许传播信息素中涉及的一对多的互动类型(Greening et al. 2015)。 自20世纪中期弗里施和他的学生的工作以来(von Frisch 1967),专注于蜜蜂信号行为的研究非常丰富。这些工作详细记录了信号在群体层面的个体影响,并讨论了(或在某些情况下建模)这种影响如何扩大到群体层面(Seeley 1995;Nieh 2010;Couvillon 2012;Kietzman and Visscher 2015)。现在,网络方法提供了一个工具箱,可以将个体和群体层面的行为联系起来。网络分析可以帮助识别信号是否有多种功能,以及不同的信号如何在功能上重叠,从而有助于理解在不同的环境背景下,尽管工人的经验有很大的不同,但蜂群级的反应如何保持稳健(Hebets et al. 2016;Hasenjager et al. 2020)。换句话说,蜜蜂群体是一个复杂的系统,信号相互作用并相互反馈以控制群体行为,而网络科学提供了一种手段,既可以理解这些以前难以理解的机制,又可以理解选择如何在一个高度社会化的群体中塑造通信系统。 参考文献 Anderson C, Ratnieks FLW (1999) Worker allocation in insect societies: coordination of nectar foragers and nectar receivers in honey bee (Apis mellifera) colonies. Behav Ecol Sociobiol 46:73–81 Barrett BJ, McElreath RL, Perry SE (2017) Pay-off-biased social learning underlies the diffusion of novel extractive foraging traditions in a wild primate. Proc Roy Soc Lond B 284:20170358 Bianconi G (2013) Statistical mechanics of multiplex networks: entropy and overlap. Phys Rev E 87:062806 Biesmeijer JC (2003a) The occurrence and context of the shaking signal in honey bees (Apis mellifera) exploiting natural food sources: shaking signal performance by honey bees. Ethology 109:1009–1020 Biesmeijer JC (2003b) The occurrence and context of tremble dancing in free-foraging honey bees (Apis mellifera). Behav Ecol Sociobiol 53:411–416 Biesmeijer JC, Seeley TD (2005) The use of waggle dance information by honey bees throughout their foraging careers. Behav Ecol Sociobiol 59:133–142 Blonder B, Wey TW, Dornhaus A, James R, Sih A (2012) Temporal dynamics and network analysis. Methods Ecol Evol 3:958–972 Boenisch F, Rosemann B, Wild B, Dormagen D, Wario F, Landgraf T (2018) Tracking all members of a honey bee colony over their lifetime using learned models of correspondence. Front Robot AI 5:35 Bonnell TR, Vilette C, Young C, Henzi SP, Barrett L (2021) Formidable females redux: male social integration into female networks and the value of dynamic multilayer networks. Curr Zool 67:49–57 Cao TT, Hyland KM, Malechuk A, Lewis LA, Schneider SS (2007) The influence of the vibration signal on worker interactions with the nest and nest mates in established and newly founded colonies of the honey bee Apis mellifera. Insectes Soc 54:144–149 Couvillon MJ (2012) The dance legacy of Karl von Frisch. Insectes Soc 59:297–306 Crall JD, Gravish N, Mounttle AM, Kocher SD, Oppenheimer RL, Pierce NE, Combes SA (2018) Spatial fidelity of workers predicts collective response to disturbance in a social insect. Nat Comms 9:1201 De Domenico M, Solé-Ribalta A, Omodei E, Gómez S, Arenas A (2015) Ranking in interconnected multilayer networks reveals versatile nodes. Nat Comms 6:6868 De Domenico M, Granell C, Porter MA, Arenas A (2016) The physics of spreading processes in multilayer networks. Nat Phys 12:901–906 Deneubourg J-L, Aron S, Goss S, Pasteels JM (1990) The self-organizing exploratory pattern of the argentine ant. J Insect Behav 3:159–168 Donaldson-Matasci MC, Dornhaus A (2012) How habitat affects the benefits of communication in collectively foraging honey bees. Behav Ecol Sociobiol 66:583–592 Donaldson-Matasci MC, DeGrandi-Hoffman G, Dornhaus A (2013) Bigger is better: honeybee colonies as distributed information-gathering systems. Anim Behav 85:585–592 Dong S, Tan K, Zhang Q, Nieh JC (2019) Playbacks of Asian honey bee stop signals demonstrate referential inhibitory communication. Anim Behav 148:29–37 Dornhaus A, Chittka L (2004) Why do honey bees dance? Behav Ecol Sociobiol 55:395–401 Dyer FC (2002) The biology of the dance language. Annu Rev Entomol 47:917–949 Easter C, Leadbeater E, Hasenjager MJ (2022) Behavioural variation among workers promotes feed-forward loops in a simulated insect colony. R Soc Open Sci 9:220120 Esch H (1964) Beitrage zum Problem der Entfernungsweisung in den Schwanzeltanzen der Honigbiene. Z Vgl Physiol 48:534–546 Farina WM, Grüter C, Díaz PC (2005) Social learning of floral odours inside the honeybee hive. Proc Roy Soc Lond B 272:1923–1928 Farina WM, Grüter C, Arenas A (2012) Olfactory information transfer during recruitment in honey bees. In: Eisenhardt D, Giurfa M (eds) C Galizia. Springer, Honeybee neurobiology and behavior. https://doi.org/10.1007/978-94-007-2099-2_8Chapter Farine DR (2018) When to choose dynamic vs static social network analysis. J Anim Ecol 87:128–138 Farine DR, Whitehead H (2015) Constructing conducting and interpreting animal social network analysis. J Anim Ecol 84:1144–1163 Finn KR, Silk MJ, Porter MA, Pinter-Wollman N (2019) The use of multilayer network analysis in animal behaviour. Anim Behav 149:7–22 Fisher DN, Pinter-Wollman N (2021) Using multilayer network analysis to explore the temporal dynamics of collective behavior. Curr Zool 67:71–80 Franz M, Nunn CL (2009) Network-based diffusion analysis: a new method for detecting social learning. Proc Roy Soc Lond B 276:1829–1836 Free JB (1965) The allocation of duties among worker honeybees. Symp Zool Soc Lon 14:39–59 Gernat T, Rao VD, Middendorf M, Dankowicz H, Goldenfeld N, Robinson GE (2018) Automated monitoring of behavior reveals bursty interaction patterns and rapid spreading dynamics in honeybee social networks. Proc Natl Acad Sci USA 115:1433–1438 Gil M, Farina W (2002) Foraging reactivation in the honeybee Apis mellifera L: factors affecting the return to known nectar sources. Naturwissenschaften 89:22–325 Gordon DM (2010) Ant encounters: interaction networks and colony behavior. Princeton University Press Gould JL (1975) Honey bee recruitment: the dance-language controversy: unambiguous experiments show that honey bees use an abstract language for communication. Science 189:685–693 Greening BR, Pinter-Wollman N, Fefferman NH (2015) Higher-order interactions: understanding the knowledge capacity of social groups using simplicial sets. Curr Zool 61:114–127 Greggers U, Koch G, Schmidt V, Dürr A, Floriou-Servou A, Piepenbrock D, Göpfert MC, Menzel R (2013) Reception and learning of electric fields in bees. Proc Roy Soc Lond B 280:20130528 Grüter C, Farina WM (2009) The honeybee waggle dance: can we follow the steps? Trends Ecol Evol 24:242–247 Grüter C, Ratnieks FLW (2011) Honeybee foragers increase the use of waggle dance information when private information becomes unrewarding. Anim Behav 81:949–954 Grüter C, Balbuena MS, Farina WM (2008) Informational conflicts created by the waggle dance. Proc Roy Soc Lond B 275:1321–1327 Grüter C, Segers FHID, Ratnieks FLW (2013) Social learning strategies in honeybee foragers: do the costs of using private information affect the use of social information? Anim Behav 85:1443–1449 Haldane JBS (1955) Aristotle’s account of bees’ “dances.” J Hell Stud 75:24–25 Hasenjager MJ, Dugatkin LA (2015) Social network analysis in behavioral ecology. Adv Study Behav 47:39–114 Hasenjager MJ, Hoppitt W, Leadbeater E (2020) Network-based diffusion analysis reveals context-specific dominance of dance communication in foraging honeybees. Nat Comms 11:625 Hasenjager MJ, Leadbeater E, Hoppitt W (2021a) Detecting and quantifying social transmission using network-based diffusion analysis. J Anim Ecol 90:8–26 Hasenjager MJ, Silk M, Fisher DN (2021b) Multilayer network analysis: new opportunities and challenges for studying animal social systems. Curr Zool 67:45–48 Hasenjager MJ, Hoppitt W, Leadbeater E (2022) Do honey bees modulate dance following according to foraging distance? Anim Behav 184:89–97 Hebets EA, Barron AB, Balakrishnan CN, Hauber ME, Mason PH, Hoke KL (2016) A systems approach to animal communication. Proc Roy Soc Lond B 283:20152889 Heinrich B (1981) The mechanisms and energetics of honeybee swarm temperature regulation. J Exp Biol 91:25–55 Hoppitt W, Boogert NJ, Laland KN (2010) Detecting social transmission in networks. J Theor Biol 263:544–555 I’Anson Price R, Dulex N, Vial N, Vincent C, Grüter C (2019) Honeybees forage more successfully without the “dance language” in challenging environments. Sci Adv 5:eaat0450 Jack-McCollough RT, Nieh JC (2015) Honeybees tune excitatory and inhibitory recruitment signalling to resource value and predation risk. Anim Behav 110:9–17 Jandt JM, Bengston S, Pinter-Wollman N, Pruitt JN, Raine NE, Dornhaus A, Sih A (2014) Behavioural syndromes and social insects: personality at multiple levels: behavioural syndromes and social insects. Biol Rev 89:48–67 Johnson DL (1967) Communication among honey bees with field experience. Anim Behav 15:487–492 Johnson BR, Nieh JC (2010) Modeling the adaptive role of negative signaling in honey bee intraspecific competition. J Insect Behav 23:459–471 Kirchner WH (1993) Vibrational signals in the tremble dance of the honeybee Apis mellifera. Behav Ecol Sociobiol 33:169–172 Kirchner WH, Lindauer M (1994) The causes of the tremble dance of the honeybee Apis mellifera. Behav Ecol Sociobiol 35:303–308 Koenig PA, Smith ML, Horowitz LH, Palmer DM, Petersen KH (2020) Artificial shaking signals in honey bee colonies elicit natural responses. Sci Rep 10:746. https://doi.org/10.1038/s41598-020-60421-8Article Krause J, James R, Franks DW, Croft DP (2015) Animal social networks. Oxford University Press, Oxford Lewis LA, Schneider SS, Degrandi-Hoffman G (2002) Factors influencing the selection of recipients by workers performing vibration signals in colonies of the honeybee Apis mellifera. Anim Behav 63:361–367 Lindauer M (1948) Über die einwirkung von duft—Und geschmacksstoffen sowie anderer faktoren auf die tänze der bienen. Z Vgl Physiol 36:391–432 Mallon E, Pratt S, Franks N (2001) Individual and collective decision-making during nest site selection by the ant Leptothorax albipennis. Behav Ecol Sociobiol 50:352–359 Mangan S, Alon U (2003) Structure and function of the feed-forward loop network motif. Proc Natl Acad Sci USA 100:11980–11985 Mersch DP, Crespi A, Keller L (2013) Tracking individuals shows spatial fidelity is a key regulator of ant social organization. Science 340:1090–1093 Michelsen A, Kirchner WH, Andersen BB, Lindauer M (1986) The tooting and quacking vibration signals of honeybee queens: a quantitative analysis. J Comp Physiol A 158:605–611 Milo R, Itzkovitz S, Kashtan N, Levitt R, Shen-Orr S, Ayzenshtat I, Sheffer M, Alon U (2004) Superfamilies of evolved and designed networks. Science 303:1538–1542 Munz T (2005) The bee battles: Karl von Frisch Adrian Wenner and the honey bee dance language controversy. J Hist Biol 38:535–570 Nandi AK, Sumana A, Bhattacharya K (2014) Social insect colony as a biological regulatory system: modelling information flow in dominance networks. J R Soc Interface 11:20140951 Naug D (2008) Structure of the social network and its influence on transmission dynamics in a honeybee colony. Behav Ecol Sociobiol 62:1719–1725 Naug D (2015) Social networks in insect colonies. In: Krause J, James R, Franks DW, Croft DP (eds) Animal social networks. Oxford University Press, Oxford, pp 160–170 Nieh JC (1993) The stop signal of honey bees: reconsidering its message. Behav Ecol Sociobiol 33:51–56 Nieh JC (1998) The honey bee shaking signal: function and design of a modulatory communication signal. Behav Ecol Sociobiol 42:23–36 Nieh JC (2010) A negative feedback signal that is triggered by peril curbs honey bee recruitment. Curr Biol 20:310–315 Nürnberger F, Steffan-Dewenter I, Härtel S (2017) Combined effects of waggle dance communication and landscape heterogeneity on nectar and pollen uptake in honey bee colonies. PeerJ 5:e3441 Okubo S, Nikkeshi A, Tanaka CS, Kimura K, Yoshiyama M, Morimoto N (2019) Forage area estimation in European honeybees (Apis mellifera) by automatic waggle decoding of videos using a generic camcorder in field apiaries. Apidologie 50:243–252 O’Shea-Wheller TA, Hunt ER, Sasaki T (2021) Functional heterogeneity in superorganisms: emerging trends and concepts. Ann Entomol Soc Am 114:562–574 Otterstatter MC, Thomson JD (2007) Contact networks and transmission of an intestinal pathogen in bumble bee (Bombus impatiens) colonies. Oecologia 154:411–421 Paffhausen BH, Petrasch J, Greggers U, Duer A, Wang Z, Menzel S, Stieber P, Haink K, Geldenhuys M, Čavojská J, Stein TA, Wutke S, Voigt A, Coburn J, Menzel R (2021) The electronic bee spy: eavesdropping on honeybee communication via electrostatic field recordings. Front Behav Neur 15:647224 Pastor KA, Seeley TD (2005) The brief piping signal of the honey bee: begging call or stop signal? Ethology 111:775–784 Pinter-Wollman N, Wollman R, Guetz A, Holmes S, Gordon DM (2011) The effect of individual variation on the structure and function of interaction networks in harvester ants. J R Soc Interface 8:1562–1573 Pinter-Wollman N, Hobson EA, Smith JE, Edelman AJ, Shizuka D, de Silva S, Waters JS, Prager SD, Sasaki T, Wittemyer G, Fewell J, McDonald DB (2014) The dynamics of animal social networks: analytical conceptual and theoretical advances. Behav Ecol 25:242–255 Ramsey M, Bencsik M, Newton MI (2018) Extensive vibrational characterisation and long-term monitoring of honeybee dorso-ventral abdominal vibration signals. Sci Rep 8:14571 Richardson TO, Gorochowski TE (2015) Beyond contact-based transmission networks: the role of spatial coincidence. J R Soc Interface 12:0150705 Richardson TO, Mullon C, Marshall JAR, Franks NR, Schlegel T (2018) The influence of the few: a stable ‘oligarchy’ controls information flow in house-hunting ants. Proc Roy Soc Lond B 285:20172726 Riley JR, Greggers U, Smith AD, Reynolds DR, Menzel R (2005) The flight paths of honeybees recruited by the waggle dance. Nature 435:205–207 Schneider SS, Lewis LA (2004) The vibration signal modulatory communication and the organization of labor in honey bees Apis mellifera. Apidologie 35:117–131 Seeley TD (1992) The tremble dance of the honey bee: message and meanings. Behav Ecol Sociobiol 31:375–383 Seeley TD (1994) Honey bee foragers as sensory units of their colonies. Behav Ecol Sociobiol 34:51–62 Seeley TD (1995) The wisdom of the hive: the social physiology of honey bee colonies. Harvard University Press Seeley TD (1998) Thoughts on information and integration in honey bee colonies. Apidologie 29:67–80 Seeley TD, Buhrman SC (1999) Group decision making in swarms of honey bees. Behav Ecol Sociobiol 45:19–31 Seeley TD, Towne WF (1992) Tactics of dance choice in honey bees: do foragers compare dances? Behav Ecol Sociobiol 30:59–69 Seeley TD, Visscher PK (2008) Sensory coding of nest-site value in honeybee swarms. J Exp Biol 211:3691–3697 Seeley TD, Kühnholz S, Weidenmüller A (1996) The honey bee’s tremble dance stimulates additional bees to function as nectar receivers. Behav Ecol Sociobiol 39:419–427 Seeley TD, Weidenmüller A, Kühnholz S (1998) The shaking signal of the honey bee informs workers to prepare for greater activity. Ethology 104:10–26 Seeley TD, Visscher PK, Schlegel T, Hogan PM, Franks NR, Marshall JAR (2012) Stop signals provide cross inhibition in collective decision-making by honeybee swarms. Science 335:108–111 Sherman G, Visscher PK (2002) Honeybee colonies achieve fitness through dancing. Nature 419:920–922 Silk MJ, Finn KR, Porter MA, Pinter-Wollman N (2018) Can multilayer networks advance animal behavior research? Trends Ecol Evol 33:376–378 Sumpter DJT (2006) The principles of collective animal behaviour. Phil Trans Roy Soc Lond B 361:5–22 Tan K, Dong S, Li X, Liu X, Wang C, Li J, Nieh JC (2016) Honey bee inhibitory signaling is tuned to threat severity and can act as a colony alarm signal. PLOS Biol 14:e1002423 Thom C (2003) The tremble dance of honey bees can be caused by hive-external foraging experience. J Exp Biol 206:2111–2116 Thom C, Gilley DC, Tautz J (2003) Worker piping in honey bees (Apis mellifera): the behavior of piping nectar foragers. Behav Ecol Sociobiol 53:199–205 Thom C, Gilley DC, Hooper J, Esch HE (2007) The scent of the waggle dance. PLoS Biol 5:e228 Vijayaraghavan VS, Noël P-A, Maoz Z, D’Souza RM (2015) Quantifying dynamical spillover in co-evolving multiplex networks. Sci Rep 5:15142 von Frisch K (1967) The dance language and orientation of bees. Harvard University Press Wario F, Wild B, Rojas R, Landgraf T (2017) Automatic detection and decoding of honey bee waggle dances. PLoS One 12:e0188626 Waters JS, Fewell JH (2012) Information processing in social insect networks. PLoS One 7:e40337 Wenner AM, Wells PH (1990) Anatomy of a controversy: the question of a “language” among bees. Columbia University Press Wenner AM, Wells PH, Johnson DL (1969) Honey bee recruitment to food sources: olfaction or language? Science 164:84–86 Wild B, Dormagen DM, Zachariae A, Smith ML, Traynor KS, Brockmann D, Couzin ID, Landgraf T (2021) Social networks predict the life and death of honey bees. Nat Comms 12:1110 Wray MK, Klein BA, Seeley TD (2012) Honey bees use social information in waggle dances more fully when foraging errors are more costly. Behav Ecol 23:125–131 Kietzman PM, Visscher PK (2015) The anti-waggle dance: use of the stop signal as negative feedback. Front Ecol Evol 3 103389/fevo201500014 原标题:《从个体到集体:基于复杂网络的方法理解蜜蜂的集体行为》 阅读原文 |
【本文地址】
今日新闻 |
点击排行 |
|
推荐新闻 |
图片新闻 |
|
专题文章 |