| 【2021年第7期 专刊】纳米材料介导微生物胞外电子传递过程的研究进展 | 您所在的位置:网站首页 › 雪娥家境 › 【2021年第7期 专刊】纳米材料介导微生物胞外电子传递过程的研究进展 |
【2021年第7期 专刊】纳米材料介导微生物胞外电子传递过程的研究进展
|
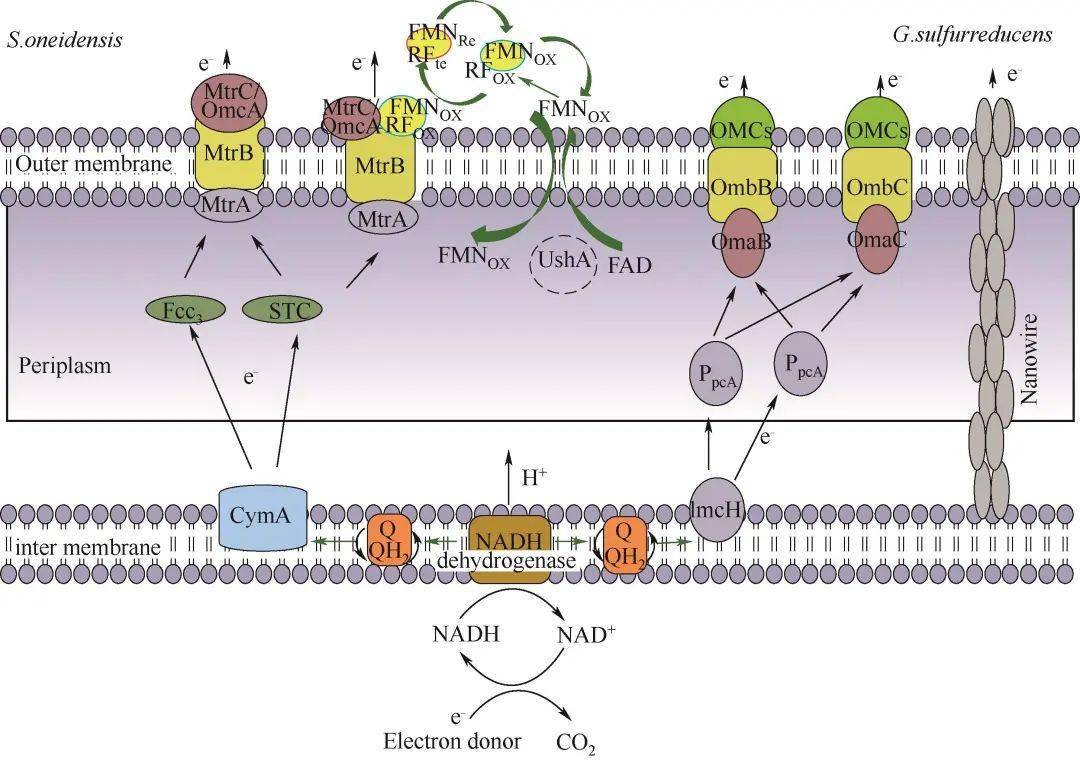
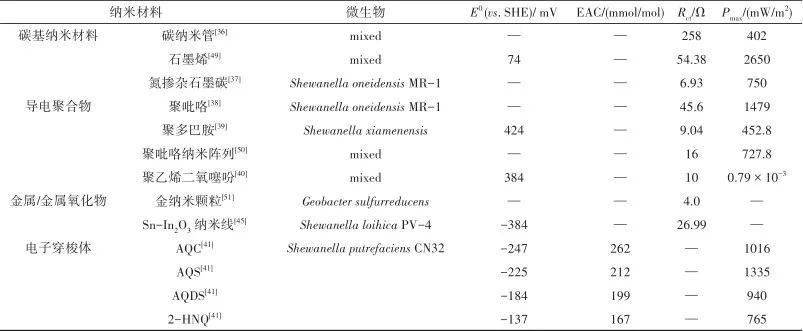
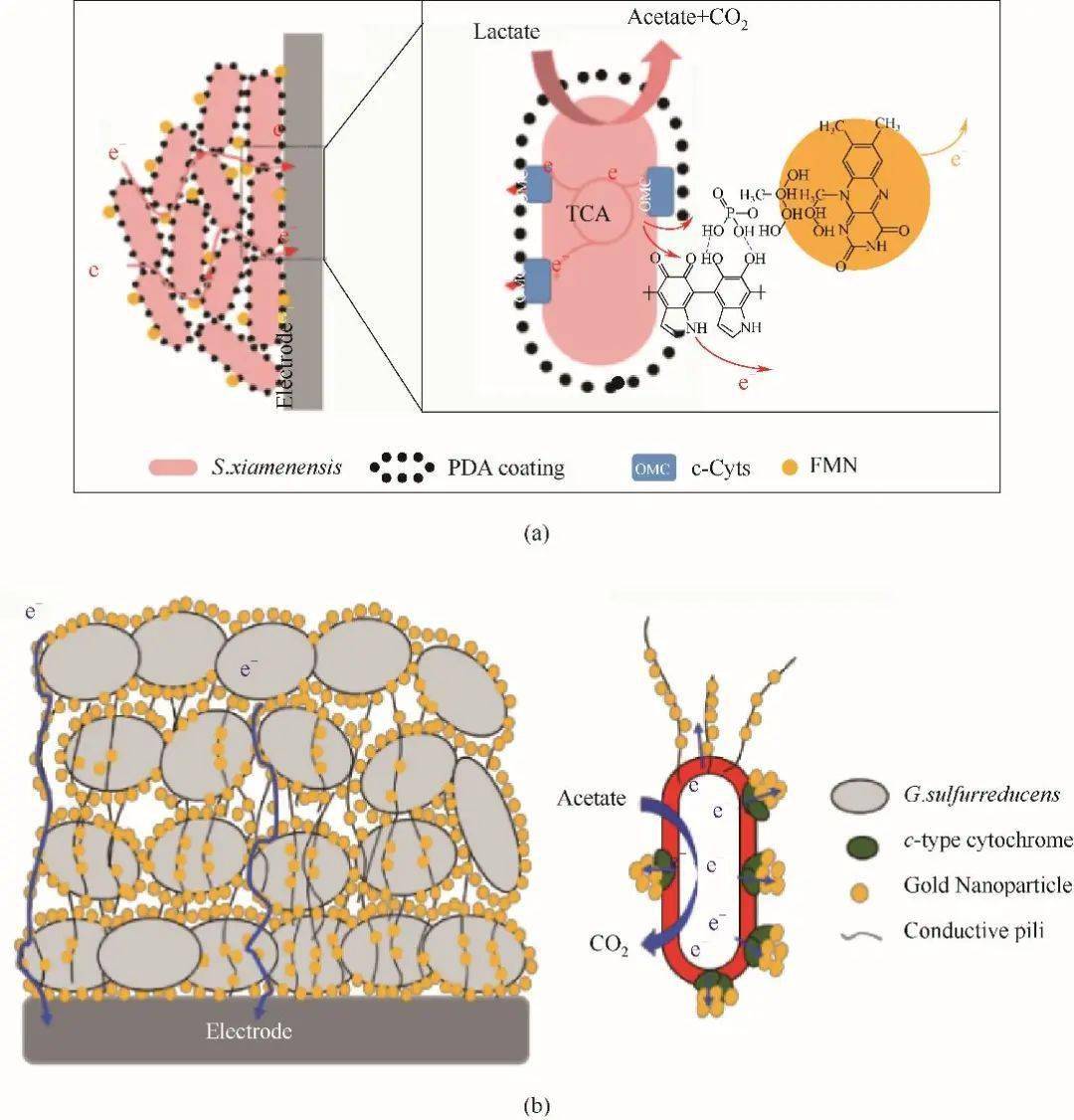
图1S. oneidensis和G. sulfurreducens胞外电子传递方式示意图[8-9 ,12]Fig.1Extracellular electron transfer model of S. oneidensisand G. sulfurreducens[8-9 ,12] 1.1 直接电子传递 1.1.1 细胞色素 c 电活性微生物可以依靠细胞胞外膜上的细胞色素 c 与胞外受体之间的直接物理接触来传递电子 [ 13] 。 S. oneidensis 中的电子传递途径被研究者们称为Mtr途径(metal reduction pathway),代谢过程中产生的电子先被细胞膜内的细胞色素CymA接受,再通过 c 型色素蛋白Fcc3和STC跨过周质空间传递至MtrCAB复合体 [ 14] 。然而,周质空间介导电子的细胞色素并不仅限于Fcc3和STC,Sturm等 [ 12] 证明同时敲除这两种细胞色素蛋白表达相关基因的突变株,在经过长时间的滞后期后仍能在柠檬酸中生长。MtrCAB复合体负责希瓦氏菌的周质空间和外膜表面的电子连接,其中MtrA和MtrC分别从周质和细胞外侧锚定在MtrB形成的孔道内。暴露于细胞外膜表面的MtrC和OmcA,作为呼吸链末端蛋白,实现胞外电子受体的还原 [ 15] 。 G. sulfurreducens 的EET模式与 S. oneidensis 类似,位于内膜与细胞质相连的MacA可以接受醌池传递出的电子,并将其转移给周质中的三亚铁红素PpcA或PpcD,跨过周质空间后,再将电子传递给外膜蛋白OMCs(包括OmcB、OmcE、OmcS及OmcZ),最终传递至胞外的固体电子受体上,完成了电子的向外传递 [ 16-17 ] 。 1.1.2 纳米导线 微生物纳米导线(microbial nanowires)是从细胞表面延伸出长达数十微米的导电鞭毛 [ 18] 。这种导电结构可以促进生物膜内微生物与固体电子受体的接触,也可以加强种间微生物电子传递。不同种微生物之间潜在的电子传递机制存在差异。研究发现,在 S. oneidensis 中存在两种类型的纳米导线,一种是基于导电的Ⅵ型菌毛,被认为可以通过依赖芳香氨基酸间电子跃迁的方式传递电子 [ 19] ,另一种由El-Naggar等 [ 20] 最近发现 S. oneidensis 外膜与周质空间可以向外延伸形成富含细胞色素 c 的膜囊泡纳管,电子通过纳管上的细胞色素 c 以跃迁方式传递。对于 G. sulfurreducens ,多数研究者也认为 G. sulfurreducens 可以表达多种纳米导线。早期研究发现 G. sulfurreducens 鞭毛上有PliA蛋白,PliA蛋白羧基端芳香族氨基酸π-π轨道的重叠,使鞭毛具有与金属相似的导电性,从而引起电子传递 [ 21] 。而最新的冷冻电镜结果表明,在 G. sulfurreducens 的表面还存在OmcS纳米导线,OmcS纳米导线中的血红素分子连续排列,空间间距3.5~6 Å(1 Å=0.1 nm),构成电子传递的通路。Pirbadian等 [ 22] 研究发现,希瓦氏菌的纳米导线是其细胞膜的外突而非蛋白微丝,与地杆菌的纳米导线有本质区别。Malvankar等 [ 23] 用静电力显微镜证实了 G.sulfurreducens 的纳米导线连接着细菌,并实现细菌间的电子传递。因此,这种优良的导电性和特殊的结构形态显示出纳米导线机制在促进远距离EET方面的潜力。 1.2 间接电子传递 间接电子传递是基于电子穿梭分子在电子供体和受体之间的扩散 [ 24-25 ] 。与整个细胞相比,这些分子具有更小的尺寸和更高的扩散,增强了它们与固体电子受体可用表面积的界面能力,从而大大提高EET效率。研究发现微生物能自分泌到细胞外具有电子传递功能的穿梭体,如黄素类、吩嗪类等 [ 26-27 ] 。Brutinel等 [ 28] 证实了 S. oneidensis 分泌黄素类物质作为电子穿梭体介导EET的可能机制,如图1所示,微生物在胞内分解有机物所产生的部分黄素腺嘌呤二核苷酸(flavin adenine dianucleotide,FAD)被转运至周质空间,然后在UshA(5'-核苷酸酶)的作用下水解为黄素单核苷酸(flavin mononucleotide,FMN)和腺苷一磷酸(adenosine monophosphate,AMP) [ 29] 。FMN可以自由穿过外膜扩散至胞外,也或者进一步水解为核黄素(riboflavin,RF)。胞外游离态的FMN ox 得到来自MtrC/OmcA的电子而被还原为FMN red ,然后再通过扩散将电子传递至胞外电子受体紧接被氧化,以此方式来介导电活性微生物与胞外受体间的电子传递过程。除了微生物自分泌的小分子,天然存在或人工合成的可溶性分子也具有传递电子的能力,如醌类、腐殖质等 [ 30-31 ] 。另外,Okamoto等 [ 32] 研究发现 S. oneidensis 分泌的黄素,除了以游离形式存在,还可以与外膜细胞色素形成细胞色素-半醌复合物进行电子传递。这种复合物的氧化还原反应速率比游离形式的黄素快10 3 ~10 5 倍。但这种结合态会随着 S. oneidensis 代谢的强弱而形成与消散,调控着EET的程度以及细胞内代谢活动。对于这种结合形式,电子如何从复合物传递至电子受体,是继续穿梭?还是直接接触电子受体表面传递?以及这个过程对微生物的重要性如何?目前还不清楚。 2 纳米材料影响EET的主要因素 电活性微生物至胞外受体的电子转移能力是影响微生物EET体系效率的关键。纳米科学的快速发展对微生物与胞外受体界面修饰从而促进EET提供了重要支撑。已有研究表明,引入纳米材料可以降低电子从电活性微生物到胞外受体的界面阻力,增强电子从电活性微生物到胞外受体的转移,改善微生物的附着和生物膜的形成,并促进特定的代谢活性 [ 33] 。纳米材料-电活性微生物界面电子传递过程受多种因素影响,如电子转移能力、氧化还原电势、表面结构与性质、生物相容性、界面相互作用、微生物种类、电子受体以及外界环境等。这里主要综述了纳米材料的电子转移能力、氧化还原电势、表面结构与性质、生物相容性以及纳米材料-微生物界面构筑对EET过程的影响。 2.1 纳米材料的电子转移能力 纳米材料能够介导电活性微生物EET与其自身电子转移能力(导电性、表面官能团等)密切相关 [ 34] 。一般来说,纳米材料的导电性越高,越可以有效地接受电子并从微生物传递至终端受体。碳基材料具有高的离域电子结构和载流子迁移率,并且碳基材料表面的氧化还原基团也具有辅助间接电子传递的效果 [ 35] ,因此常被应用于增强生物膜的导电性进而促进EET。Liang等 [ 36] 考察了碳纳米管、活性炭和柔性石墨分别作为微生物燃料电池阳极对 S. oneidensis MR-1产电性能影响,其中以碳纳米管作为阳极的MFC的功率密度最高,内阻最低,分别为402 mW/m 2 和263 Ω。研究结果表明,相比于其他两种碳基材料,碳纳米管能够有效地降低MFC的阳极内阻和欧姆内阻。循环伏安法分析表明,碳纳米管的介导使微生物EET动力学增强,包括 S. oneidensis MR-1的外膜细胞色素OmcA的氧化还原活性提高。除了利用纳米材料本身性质外,还可以通过引入表面缺陷和杂原子掺杂来调节纳米材料的表面电荷转移性质。如You等 [ 37] 使用氮原子掺杂打破碳纳米管的化学惰性,在表面形成更多的缺陷,实验剖析证实多孔纳米碳材料表面基团与产电菌间的相互作用,有利于提高细菌胞外直接电子传递,进而改善MFC性能。 除了碳基纳米材料,研究人员也曾利用导电聚合物(如聚吡咯 [ 38] 、聚多巴胺 [ 39] 、聚乙烯二氧噻吩 [ 40] 等)介导EET过程,一方面降低了微生物至电极受体之间的传质内阻( R ct ),以获得更高的MFC电流输出;另一方面同时改善了生物相容性,以达到MFC长期有效运行。金属/金属氧化物具有更高的导电能力或易调控的氧化还原电势,也被引入介导EET过程。为了单一评估电子转移能力对EET的影响,Li等 [ 41] 将Fe(Ⅲ)作为末端电子受体,不同醌类作为介导电子转移的氧化还原中间体,探究氧化还原中间体的电子转移能力与EET速率的相关性。研究发现Fe(Ⅲ)还原速率与中间体的电子转移能力呈很好的线性正相关关系,相关系数高达0.933,表明纳米材料介导的微生物EET过程与纳米材料的电子转移能力密不可分;同时研究者发现将电极作为电子受体时,EET速率与电子转移能力的相关系数只有0.801,这表明了纳米材料的电子转移能力对EET的影响,不仅是纳米材料本身性质,也同时受其他因素影响,如电子受体等。 2.2 纳米材料的氧化还原电势 电活性微生物通过内源或外源电子穿梭体促进EET,主要是因为这些穿梭体具有合适的氧化还原电势,能够接受细胞色素 c 的电子变成还原态,再将其传递至胞外受体随之被氧化。纳米材料与微生物外膜、电子受体之间的电势差,决定着电子传递反应的驱动力。Liu等 [ 42] 研究发现在 G. metallireducens 和 G. sulfurreducens 共培养体系中,Anthraquinone-2, 6-disulfonate(AQDS)能增强乙醇氧化和琥珀酸还原,但其无法介导 G. metallireducens 和 Methanosarcina barkeri 互营促进甲烷产生过程,主要因素可能是氧化还原电势。AQDS/AHQDS氧化还原电势为 E 0 = - 184 mV,可驱使延胡索酸的还原,而将CO 2 还原成CH 4 所需的电势为 E 0 (CO 2 /CH 4 )= - 240 mV,高于AQDS/AHQDS氧化还原电势。Li等 [ 43] 研究发现,将可溶的萘醌负载在氧化石墨烯上制备成固态的醌类化合物,能很好地促进微生物的铬还原能力。因此,与可溶性穿梭体类似,具有合理氧化还原电势的固态纳米材料也可以作为氧化还原介体介导微生物的电子传递。 一般来说,微生物由呼吸作用获取的能力与纳米材料的中点氧化还原电势具有相关性 [ 27] 。理论上,纳米材料的氧化还原电势应介于两个氧化还原的半反应之间 [ 44] 。纳米材料的氧化还原电势不应低于终端电子受体的电势,否则纳米材料无法转移电子;同理,若氧化还原电势比电子受体高太多,则该反应无法顺利进行 [ 44] 。Bian等 [ 45] 通过Sn掺杂In 2 O 3 纳米线(SINWs)调节其氧化还原电势到所需的范围,以降低细菌-电极界面的能垒,与未修饰纳米线的平面FTO电极相比,SINW修饰后可以使电流提高60倍。微生物外膜的细胞色素氧化还原电势位于-0.2 V,希瓦氏菌分泌的黄素氧化还原电势为 -0.4 V,合成Sn掺杂的In 2 O 3 纳米线的氧化还原电势位于-0.57 V,在+ 0.2 V的偏压下,细胞外膜传递的电子可以被黄素的氧化态接收,而合成的纳米线可以接受黄素还原态的电子并最后转移到电极上。黄素/丙二酸的外源添加只增强/阻碍这一间接电子传递过程,也进一步证实了合成的纳米线可促进间接电子传递过程。基于类似的策略,其他具有适当氧化还原电势的纳米材料,如α-Fe 2 O 3 、γ-Fe 2 O 3 和Fe 3 O 4 可以匹配细菌OMCs的能级并缩小电荷转移间隙,也被开发为可以有效介导EET的纳米材料 [ 46-47 ] 。O'Loughlin [ 48] 采用多种纳米材料介导 S. putrefaciens CN32还原纤铁矿,研究显示纳米材料在表观电势为-254 ~ + 11 mV条件下与Fe还原速率具有较好的相关性,其中 r 2 高达0.973。因此,热力学因素氧化还原电势不仅决定着纳米材料介导EET的顺利进行,还对纳米材料介导EET的速率具有很大影响。表1总结了部分纳米材料的氧化还原电势和电子转移能力。 表1 纳米材料介导微生物EETTable 1 Nanomaterials mediated microbial extracellular electron transfer
注:E 0 为标准氧化还原电位;EAC为电子接受能力(electron accepting capacity); R ct 为传质内阻; P max 为最大输出功率密度;AQC为蒽醌-2-羧酸(anthraquinone-2-carboxylic acid);AQS为2-蒽醌磺酸(2-anthraquinonesulfonic acid);AQDS为蒽醌-2,6-二磺酸二钠(anthraquinone-2,6-disulfonic acid disodium);2-HNQ为2-羟基-1,4-萘醌(2-hydroxy-1,4-naphthoquinone)。 2.3 纳米材料的表面结构与性质 纳米材料的微纳拓扑结构能有效直接影响微生物的黏附、增殖以及分化等生物学行为 [ 52] 。比如纳米颗粒、纳米管、纳米线、纳米纤维和纳米片,不同的形貌会造成电子传导距离和接触活性位点面积的不同,因此会影响电子传递效率。Xie等 [ 53] 使用溶剂诱导平面基底上的碳纳米管薄膜先溶胀后收缩,形成三维的褶皱结构,研究发现经处理后形成的拓扑结构会促进细胞和纳米材料之间的相互作用从而影响细胞功能。起皱的碳纳米管膜利于微生物的附着、生长和电子转移,这种表面结构构建的阳极,使微生物燃料电池的输出电流提高了2.6倍。三维的孔道结构无论是在化学催化领域还是生物催化领域,都是目前纳米材料结构设计的热点方向,主要是因为这种结构不仅允许微生物的内部定殖,还有利于底物和介质的畅通运输,保证了生物膜中细胞的高生存能力和高效的EET [ 54] 。另外,层级有序的三维连通孔道可提供多方向、高通量离子运输通道,有助于提升表/界面反应动力学。Holder等 [ 55] 采用分凝成冰诱导的自组装技术制备了3D壳聚糖/真空剥离石墨烯复合材料并将其用于生物电化学系统。证明了大孔和中微孔的协同作用,其中相互交联的大孔可以提供细菌进入材料内部和缓冲离子进而缩短扩散距离,薄壁上的中孔以及由于石墨烯堆垛而形成的微孔提高了接受电子的面积。一些特殊巧妙的结构设计也被开发用于增强EET,如Wang等 [ 50] 使用电化学聚合法制备出了带有吸盘结构聚吡咯纳米线阵列,当其用于微生物燃料电池的阳极时,阵列上附着的细菌在新陈代谢过程会消耗阵列内管的氧气导致形成真空态,从而促使电极与细菌的强烈吸力,加快它们的相互作用,进而驱使微生物基于细胞色素 c 的直接电子传递。这种结构设计的微生物燃料电池功率密度高达728 mW/m 2 ,未来有望通过优化阵列的尺寸进一步提高电池性能。 纳米材料的表面性质(亲水性或者疏水性)也会对细胞行为产生影响。Ding等 [ 56] 研究发现纳米材料表面的湿润性极大地影响了 Shewanella loihica PV-4的EET能力,该菌在亲水性的电子表面的EET能力比在疏水性表面高5倍。研究表明,纳米材料表面的湿润性会影响胞外膜细胞色素的氧化还原态,胞外膜细胞色素为还原态时更利于外膜细胞色素与黄素结合,使胞内传递出来的电子能以黄素-细胞色素辅因子的形式进行传递,结合作用下EET效率比黄素单独作用快10 3 ~10 5 倍。 2.4 纳米材料的生物相容性 使用纳米材料与细菌结合,容易对细胞产生毒害作用,限制了微生物的催化效率。纳米材料对细胞的毒害有直接物理破坏,或者产生活性氧自由基间接破坏 [ 57] 。有研究报道石墨烯是一种具有良好生物相容性的碳基材料,但也有研究发现石墨烯具有一定的生物毒性,其尺状的边缘容易损害细胞膜,从而影响细胞的活性 [ 58] 。Li等 [ 43] 将石墨烯与聚乙烯醇混合,通过真空抽滤制成薄膜用于铬还原,不仅增加生物相容性,而且制膜可循环利用,利于经济效益。纳米材料的生物相容性并不绝对,与其形状大小、表面电荷、官能团及添加量密切相关 [ 59] 。另外,研究也发现同一纳米材料作用于不同种微生物时会呈现出完全迥异的生物毒性。 除了降低纳米材料的生物毒性外,还可以开发具有保护细胞活性的纳米材料与结构设计。Ji等 [ 60] 设计合成的MOF(metal-organic framework),能与 Morella thermoacetica 表面的壁磷酸结合形成薄膜包覆结构,细胞表面的MOF可以催化降解氧应激产生的活性氧自由基,降低了这种厌氧微生物在有氧条件下的5倍死亡率。半导体材料如CdS [ 61] 、ZnS [ 62] 、InP [ 63] 等也被广泛用于微生物系统,它不仅可以利用半导体材料优异的光吸收性能,而且通过微生物和半导体的紧密接触可以降低电子转移的电阻,实现将太阳能高效转化为电能/化学能。光激发下CdS空穴产生的活性氧物种(·OH、O 2 - ·和H 2 O 2 )对细菌会产生一定的毒性并影响其生物活性,进而限制微生物的催化活性。针对这一问题,Chen等 [ 64] 通过简单快速的方法在CdS表面修饰Mn 3 O 4 纳米酶,Mn 3 O 4 对CdS产生的活性氧物种有及时分解和物理分离的作用,原位修饰Mn 3 O 4 后·OH、O 2 - ·和H 2 O 2 的浓度分别减少了90%、77.6%和26%,氮还原提高了28%,N 2 O的生成量减少了78%。解决纳米材料与微生物作用的生物相容性问题,进而有效地增强微生物的催化活性。 2.5 纳米材料-微生物界面构筑 除了考虑纳米材料本身的性质与结构外,纳米材料与微生物相互作用的界面也决定二者间的能量和电荷传递 [ 65] 。总结以往的纳米材料与电活性微生物界面构筑的研究进展来看,从最早期研究者使用纳米材料对电子受体(如最常用的电极)进行修饰,电活性菌自然生长在纳米材料表面;到后面发展了一种新策略,纳米材料与微生物混合形成杂化的生物膜;为了更充分利用细菌胞外膜上的每一个活性位点,使用纳米材料原位修饰在细胞表面;紧接着,为了使纳米材料与微生物作用更紧密一步,将微小的纳米材料插入到细胞膜或周质空间,与细胞膜上的电子链的蛋白进一步作用;最后到近期,将超小的纳米材料引入到细胞内部,拓展了界面构筑的新思路。纳米材料介导微生物胞外电子传递的过程,通常同时受多种因素影响,对于纳米材料的选取、构筑方式,一般都以研究目的或实际应用进行兼容考虑。因此,接下来选取纳米材料-微生物界面构筑这个角度,阐述界面构筑对EET的增强机理以及优缺点,为具有类似功能的纳米材料提供构筑思路。 3 纳米材料-电活性微生物界面构筑及胞外电子传递的研究进展 3.1 电子受体表面修饰的电子传递 自然条件下电活性微生物在固体胞外受体表面形成的生物膜薄且导电性差,为此很多研究团队开发了一系列的功能纳米材料以改善细菌和胞外受体的相互作用。Li等 [ 66] 使用GO/PVA复合膜作为外源电子中介体,介导 Shewanella EET用于Cr(Ⅵ)还原体系,使还原效率提高了3.13倍[图2(a)]。研究发现,在还原过程中,GO/PVA膜不仅作为氧化还原中介体传递电子,GO/PVA膜的添加还会影响细胞的生理变化,提高电子传递链上的细胞色素 c 相关基因表达量。分别提高这些相关基因( mtrA 、 mtrB 、 mtrC 、 mtrD 、 mtrF 和 omcA )的表达量后发现,基因表达水平的提升能显著促进Cr(Ⅵ)的还原速率。Zhao等 [ 67] 通过在负载石墨烯的阳极表面修饰一层离子液体,带负电荷的阳极表面变为带正电荷。由于静电作用,经离子液体修饰的电极可以紧密地吸引带负电的希瓦氏菌,增强且加速了细菌的附着,其输出的生物电流提高了7倍。虽然电子受体表面修饰对微生物的远距离传输作用甚微,但可以有效缩短电极表面生物膜的形成时间,在实际应用中具有一定的研究价值。Du等 [ 68] 使用聚多巴胺对电极表面进行修饰,聚多巴胺改善了电极表面的亲水性,缩短电池的启动时间,经聚多巴胺修饰后形成的生物膜具有长期稳定性和耐久性,并且制备简单可以很好地放大规模。近年来,考虑到电子受体修饰是通过胞外膜细胞色素 c 和纳米导线的直接电子传递来直观有效地增强EET性能,诸多研究团队致力于开发制备3D大孔结构,增加电极表面电活性细菌的载量。如Fang等 [ 69] 设计了一种反蛋白石三维结构的锡化铟电极[图2(b)],这种电极结构有着相互连接且均匀的大孔隙(8~10 μm),细菌很容易进入到电极内部并富集生长,这样的孔结构利于电极结构内部与外界的物质传递,极大地增强细菌通过胞外膜细胞色素 c 的直接电子传递,产生的电流可达3 mA/cm 2 。此外,Zou等 [ 70] 通过高温碳化细菌纤维素制备了3D复合多孔碳纳米纤维气凝胶,通过调节碳化温度巧妙地调控其纳米孔结构(< 10 nm)。研究表明,随着碳化温度的升高而使得碳纤维气凝胶中微孔逐渐转换为介孔,通过电子穿梭体传递电子的能力增强,介孔不但增加了3D多孔碳纤维电极中电子穿梭体可利用的表面积,并且由于其孔内凹凸不平的表面和适当的扭曲结构克服了黄素分子(RF、FMN)的空间位阻,从而实现高效的两电子传递。因此,目前修饰电子受体的策略已经非常多样化,是比较容易操作和实现的,且在放大系统中也较稳定。
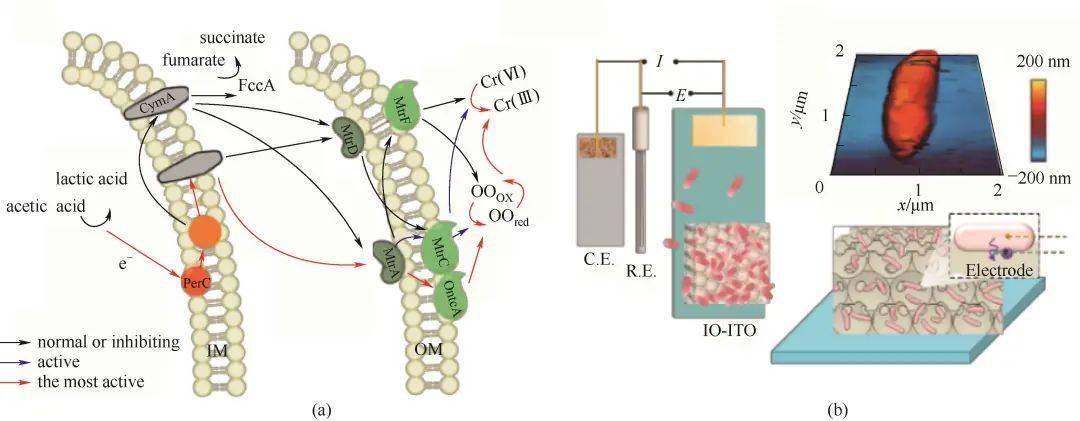
图2氧化石墨烯介导S. xiamenensis还原Cr(Ⅵ)的胞外电子传递路径图(a)[66];IO-ITO|G.sulfurreducens电极组装成三电极体系,G.sulfurreducens置于硅片上的原子力显微镜图像,G.sulfurreducens在IO-ITO骨架上形成生物杂化电极的示意图(b)[69]Fig.2The pathway diagram of graphene oxide films mediates extracellular electron transfer of S. xiamenensisto reduce Cr(Ⅵ) (a)[66]; An IO-ITO|G.sulfurreducenselectrode is assembled into a three-electrode system; Atomic force microscopy (AFM) image of G. sulfurreducenson a silicon wafer; Schematic representation of a biohybrid electrode where G. sulfurreducenscolonized on the IO-ITO scaffold (b)[69] 3.2 生物膜杂化的电子传递 微生物在自然条件下形成的生物膜,电子受体表面的负载量有限,且不利于生物膜内的细菌远距离传输。为了解决这一问题,用导电材料与微生物混合,形成电活性杂化生物膜可以加强电活性微生物远距离电子传输效率。Yong等 [ 71] 通过将氧化石墨烯(GO)添加到MFC阳极中[图3(A)],有效提高 S. oneidensis MR-1的EET效率。研究发现细菌会将与其紧密接触的GO还原为还原型石墨烯(rGO),rGO自组装成三维大孔网络结构,大量的细菌黏附在网络结构中,形成细菌-rGO导电网络的杂化生物膜,最终提高了细胞与电极间的双向电子传递,其中氧化电流(电子从细菌转移到阳极)增强了25倍,还原电流(电子从阴极转移到细菌)提高了74倍。另一项研究报道 Shewanella PV-4利用溶液中的游离态离子通过生物矿化作用形成FeS纳米颗粒,混合在微生物之间的FeS纳米能介导远距离细胞外电子转移和连接空间离散的氧化还原环境 [ 46] 。这不仅导致了生物矿化以形成纳米颗粒的发展,而且合成的生物纳米颗粒进而提高了微生物的EET效率。此外,Ma等 [ 72] 研究发现纳米材料还可以促进微生物种间的直接电子传递,促使电活性微生物直接通过微生物之间的物理性接触和纳米导线的生成,通过添加TiO 2 不仅提高了反应体系的导电性,还明显改变了反应体系中的微生物群落结构,介导并促进互营菌与产甲烷菌间种间直接电子传递,从而加速了厌氧产甲烷过程[图3(B)]。可以看出,生物膜杂化结构能有效改善微生物的远距离传输,并且在实际体系中具有重要的研究价值。
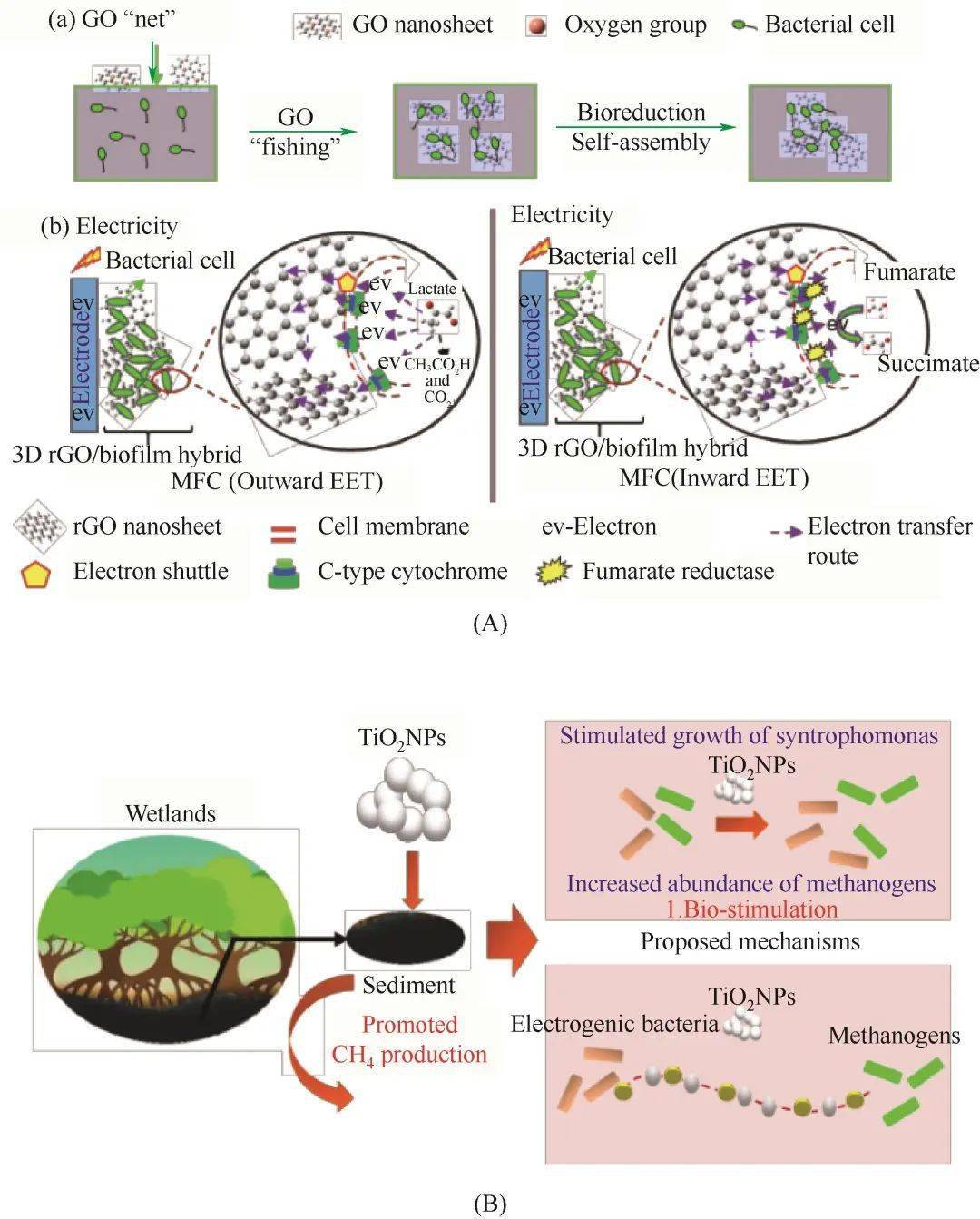
图3(A)三维多孔的还原氧化石墨烯/细菌杂交生物膜的自组装过程(a)及提出的双向EET机制(b)[71];(B)二氧化钛介导微生物共生产甲烷的原理图[72]Fig.3(A) Self-assembly of the 3D microporous rGO/bacteria hybrid biofilm by a fishing process (a), and the proposed mechanism of bidirectional EET (b)[71]; (B) Schematic diagram of TiO2-mediated methane production by microbial syntrophism[72] 3.3 细胞表面修饰的电子传递 用纳米材料制备电活性杂化生物膜虽然可提高微生物的EET速率,但是由于纳米材料不能与生物膜中的所有微生物完全接触并且与微生物的活性位点存在一定的空间位阻,导致EET效率降低。因此,用导电材料对单个细菌进行修饰是一种解决方案。如Liu等 [ 39] 采用原位聚合的方法,将聚多巴胺(PDA)这种具有醌/半醌结构的氧化还原介质 [ 73] 修饰在细菌表面,在不影响细胞生存能力的情况下明显提高了希瓦氏菌的电活性,显著改善了微生物与电极之间的相互作用[图4(a)]。其中,在以乳酸为电子供体时,修饰有PDA的希瓦氏菌的EET能力得到了明显提高,其催化的MFCs的最大输出功率密度提高了5倍。研究表明,细胞外膜上的 c 型细胞色素能够通过细胞表面形成的氧化还原中介体PDA涂层进行电子传递,即使细菌远离电极,也可以通过PDA将产生的电子有效地传递到电极,从而增强了细胞色素 c 介导的EET。更重要的是,细菌表面PDA涂层还可以吸附细菌自分泌的电子穿梭 体——黄素增强间接电子传递,从而在细胞表面形成了多重传递电子路径。Chen等 [ 51] 报道了一种简单、高效的提高地杆菌 Geobacter sulfurreducens 生物发电能力的方法[图4(b)]。通过生物矿化作用原位在细菌表面形成Au纳米粒子,使得该菌的胞外电子转移能力增强。在生物电化学系统中,表面负载Au的细胞与未负载相比,最大电流密度提升了40%,有机质(乙酸)的去除增强了2.2倍。这些研究表明,用纳米材料对单个微生物细胞进行修饰也能显著加强微生物EET能力,提高细胞与电极间电子传递效率。但由于细胞的分化生长可能会影响系统的稳定,因此比较适用于小型化如生物传感等应用。
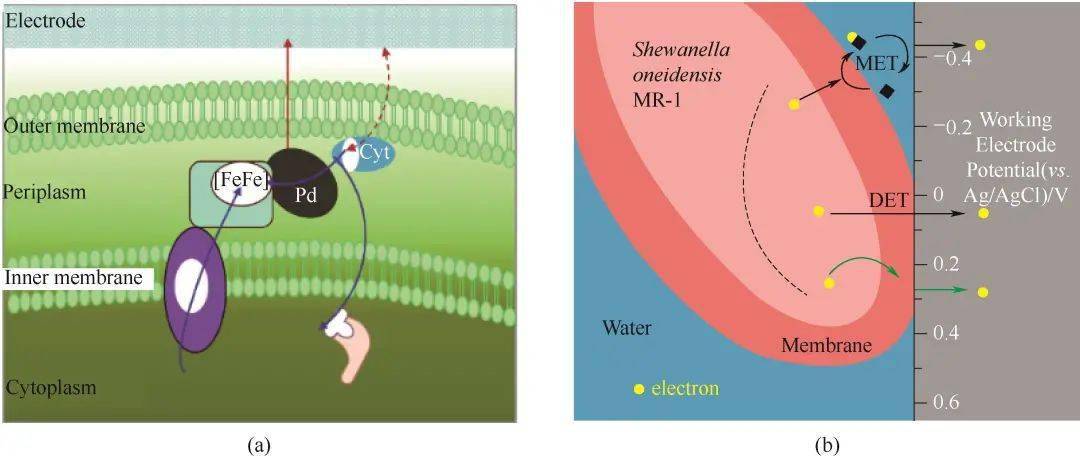
图4在活细胞表面修饰聚多巴胺以增强细胞外电子传递的机制示意图(a)[39];原位制备的金纳米颗粒增强BES电流的机理图(b)[51]Fig.4Proposed mechanism of coating polydopamine (PDA) on individual live cells to enhance extracellular electron transfer(a)[39]; Proposed mechanism of a current enhanced of BES with in situAu-NP fabrication(b)[51] 3.4 跨细胞膜的电子传递 有报道称,适当尺度的金属和低聚物纳米材料可以结合进入细胞的细胞膜,突破EET的第一道屏障进而加速细胞跨膜的电子传递过程。Wu等 [ 74] 通过以脱硫弧菌 Desulfovibrio desulfuricans 为模型生物来研究膜结合生物合成的Pd纳米颗粒对该菌EET过程的影响[图5(a)]。研究表明了Pd纳米颗粒可出现于细胞周质空间并代替细胞色素蛋白传递电子,其可沿着天然酶介导的EET途径来加快电子转移。生物合成的纳米材料优势在于它们可以对氧化还原蛋白具有较高的识别能力,并能充分地插入膜中,这是设计合成的纳米材料无法轻易实现的。共轭聚合物不仅具有高导电性,并且该分子的π-共轭芳族骨架可以使电荷离域化,而侧链的离子基团则赋予了其良好的水溶性,这种两亲分子可以与细胞膜发生物理相互作用或直接穿过细胞膜插入细胞中,从而可以使细胞进行直接EET [ 75] 。Kirchhofer等 [ 76] 通过用二茂铁基的共轭聚合物分子——DSFO+与希瓦氏菌作用,该方法极大地改善了细菌和电极之间的EET,DSFO+首先通过静电力与希瓦氏菌相互作用,然后通过疏水作用插入细菌细胞壁,从而与细胞色素 c 紧密接触,调节希瓦氏菌跨膜的电子传递过程[图5(b)]。因此,DSFO+可以直接催化跨膜的呼吸电流,这种策略为加强纳米材料和微生物的相互作用更推进一步。然而,目前仍然无法明确低聚物促进微生物EET过程的具体机制,包括促进细胞膜通透性、细菌附着和加快电子传递速率等方面都需要在未来进行研究。
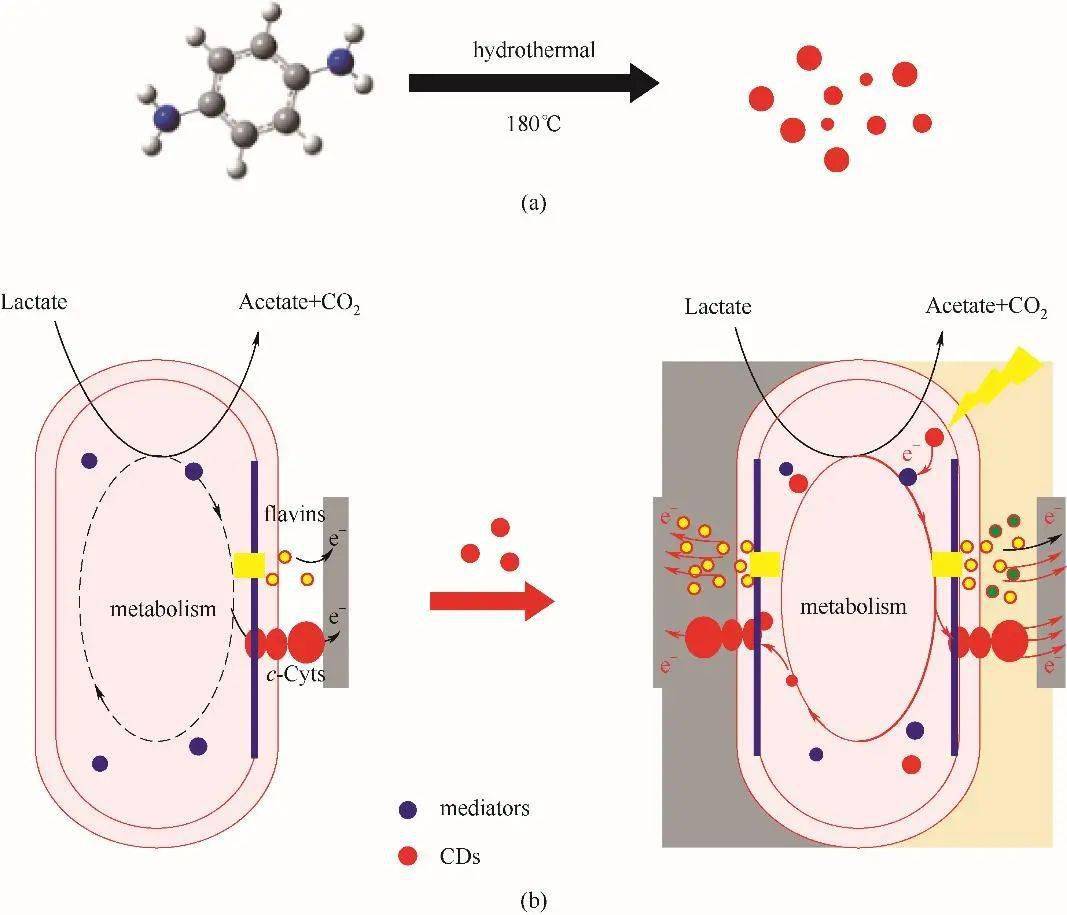
图5细胞和电极之间胞外电子传递链的假设途径(a)[74];膜插层共轭氧化还原分子(DSFO+)催化跨膜电子转移(b)[76]Fig.5Hypothesized pathways for the extracellular electron-transfer chain between the cell and the electrode(a)[74]; Membrane-intercalating conjugated redox molecule (DSFO+) catalyzes transmembrane electron transfer(b)[76] 3.5 细胞内化的电子传递 纳米材料的细胞内在化可引起广泛的生物反应,也可激活细胞内信号通路 [ 77] 。由于驱动EET通路的生物催化电子池位于细胞质内,纳米材料穿过细胞膜进入细胞质可能会产生额外的相互作用,影响电子传递、细菌生理和细胞功能。近期,Liu等 [ 78] 通过一步水热法合成直径约为7 nm的碳量子点(CDs),仅通过简单添加的方式,合成的碳量子点可以被希瓦氏菌内化进胞内,且具有优良的生物相容性(图6)。在黑暗条件下,内化的碳量子点不仅增强了细菌的电导性,而且还刺激了细菌的生理反应使其分泌大量的黄素。另外,在模拟的太阳光照射下,胞内的碳量子点受光激发产生的电子进入到胞内代谢中,驱动了希瓦氏菌的厌氧呼吸进而增强了EET。该研究工作不仅为改善EET性能提供了一种简便且有效的策略,更为重要的是为探究碳基半导体与微生物的相互作用奠定了基本认识。但关于其生理应激的反应机理和电子转移途径的阐明目前仍然是一个挑战。另外,除了单独的生物电催化,引入其他驱动力如太阳能来主动驱动细胞内的反应,实现多维能量转换也是一种有效策略。
图6对苯二胺水热法合成具有红色荧光的CDs(a); S. xiamenensis在有CDs (左)和无CDs(右:暗/光)情况下的假设电子转移模式示意图(b)[78]Fig.6Synthesis of CDs with red fluorescence using hydrothemal treatment of p-phenylenediamine (a); Schematic illustration of the hypothesized electron transfer pattern in the absence (left) and presence (right:dark/light) of CDs for cells of S. xiamenensis(b)[78] 4 展 望 EET是近年来土壤、生物、能源和环保等研究领域的热点,EET效率的提高是其能否从实验室走向实际应用的核心。纳米材料与电活性微生物的结合能够有效增强EET效率。微生物的自身特性、纳米材料的性质以及微生物细胞与纳米材料的结合形式都会对纳米材料与微生物的界面相互作用起决定性作用。因此,后续的研究当致力于以下几个方面:(1)从电活性微生物本身出发,在引入纳米材料强化的同时,借助分子生物学、生物化学、电化学、物理学等方法进行机理解析,不仅能够明确与电活性细菌EET相关的组分及传递途径,还可以为使用基因工程手段进一步强化作指导。(2)从纳米材料制备角度来看,用于增强EET过程的纳米材料需要特定的设计来匹配不同的EET途径,并解决纳米材料自身独特的理化性质与微生物催化更好的结合与兼容性问题,在材料的生物相容性和微生物的化学相容性之间探索出平衡点。另外,新材料的合成也迫切需要开发,如具有特殊结构的材料、仿生材料和天然材料等。(3)纳米材料与细胞膜结合或进入到胞内的策略,目前仅包括低聚物和部分金属/半导体的改性,这表明在材料设计挖掘方面还有很大的发展空间。另外,纳米材料与细胞相互作用的机制也亟待研究。(4)根据微生物的特性和最终的应用需求选择合适的材料体系,包括纳米材料-电活性微生物界面构筑的策略,可以从这些策略的适用性和局限性进行拓展以及改善。(5)目前的方法主要通过电化学技术观察纳米材料-电活性微生物整体电流或输出功率来评估复合材料的性能。但是,传统电化学技术不能揭示EET的亚种群差异,不能将观察到的电化学电流与单个修饰材料的细胞联系起来,忽略了生物电化学系统活动的潜在复杂空间模式,无法明确区分在整体电化学性能变化背后的机理。后续在控制并成像单细胞水平的纳米材料-电活性微生物/电极相互作用的工作也会推动对EET的进一步理解。 Progress in nanomaterials mediated microbial extracellular electron transfer LIU Shurui WU Xue'eWANG Yuanpeng Abstract: The process of microbial extracellular electron transfer (EET) is widespread in nature and has broad application prospects in energy utilization and environmental remediation. However, inefficient electron transfer has always been a key bottleneck in practical applications. Nanomaterials have unique properties such as surface effect, volume effect, quantum size, and macro-quantum tunneling effect. The combination of nanomaterials and electroactive microorganisms can achieve complementary advantages, which can shorten the charge transfer path and increase the rate of EET. This review introduces the pathways of EET, as well as the factors affecting the interface EET such as the electron transfer ability, redox potential, surface structure and biocompatibility of nanomaterials, with a focus on various strategies for constructing the interface between nanomaterials and electroactive microorganisms, and the applicability and limitations of these strategies are summarized. Finally, the future research direction of nanomaterials to enhance electroactive microorganism EET is prospected. Keywords: electroactive microorganism;extracellular electron transfer;nanomaterials;interface;biocatalysis 第一作者:刘姝睿(1992—),女,博士研究生,[email protected] 通信作者:王远鹏(1979—),男,博士,教授,[email protected] 返回搜狐,查看更多 |
【本文地址】