| 文献阅读 | 您所在的位置:网站首页 › 葡萄树的一生之注释 › 文献阅读 |
文献阅读
|
Liang, Z., Duan, S., Sheng, J. et al. Whole-genome resequencing of 472 Vitis accessions for grapevine diversity and demographic history analyses. Nat Commun 10, 1190 (2019). https://doi.org/10.1038/s41467-019-09135-8 在基因组水平上了解葡萄的种类对于葡萄品种的改良非常重要。本文作者对472种葡萄进行了重测序,涵盖了来自广泛地理分布的60个现存葡萄树中的48个物种。这有助于确定最近驯化的葡萄中有效种群数量的急剧膨胀和收缩。此外,作者发现候选基因与重要的农艺性状(如浆果形状和芳香化合物)之间存在关联。这些结果证明了重测序数据的资源价值,以阐明葡萄品种的进化生物学并为葡萄遗传改良提供目标。 在葡萄研究中,除了常用的 10-20K 基因型芯片外,葡萄研究中共仅有最近的36个样本得到了全基因组重测序。基因组资源的缺乏阻碍了对葡萄的过去历史和现在的性状改良的研究。本次作者测定的种质包括欧洲、东亚、北美等地的190个野生种、220个栽培种和109个杂交种。 目录SNP和基因组结构变异葡萄系统发育分析葡萄种群结构分析连锁不平衡分析WEU和CEU葡萄树的群体历史系谱分析WEU和CEU中的选择信号 SNP和基因组结构变异首先是测序结果的统计。共产生了约4.1 Tb的全基因组测序数据,平均深度约为15.5x,对 V. vinifera reference genome 的比对率为 97.4 ± 4.6%,估计错误率为 0.02 ± 0.01%。主要accession全部染色体的基因组覆盖率超过80%。
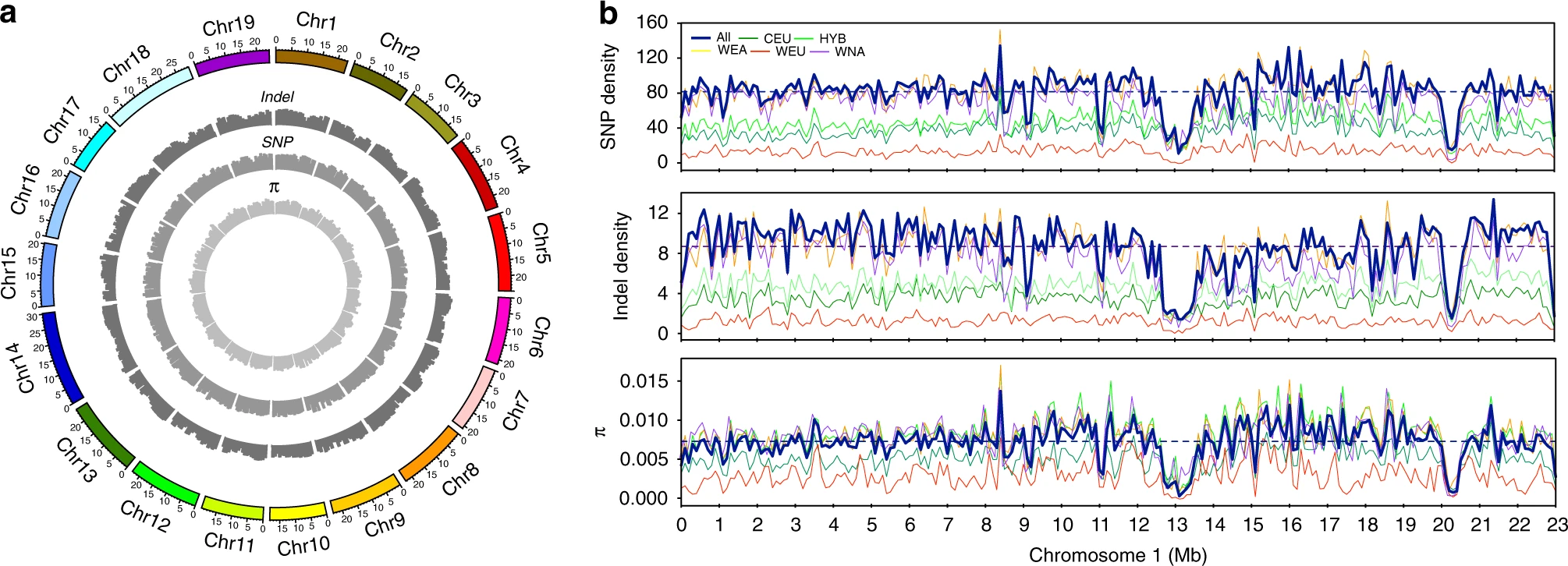
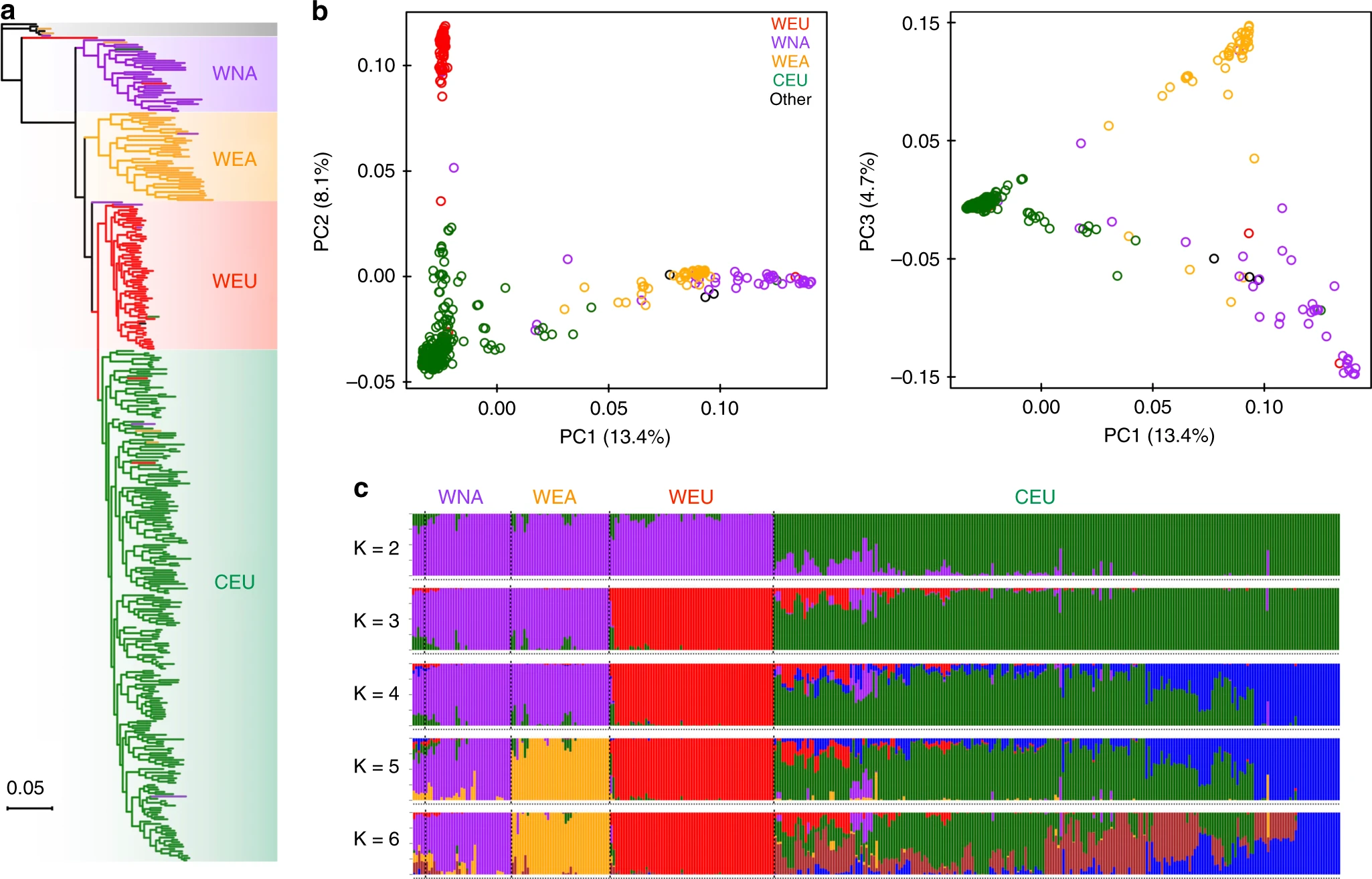
图1 在472个葡萄属种质中鉴定出的基因组变异的摘要。a 葡萄基因组所有染色体上SNP密度、indel密度和总遗传多样性(π)的Circos图表示,缩放至Mb大小。b 连续100 kb窗口的葡萄基因组1号染色体上SNP密度,indel密度和总遗传多样性图。虚线代表整个基因组的平均值。WNA (紫色),WEA(黄色),WEU(红),CEU(深绿色),和HYB(浅绿色)代表北美野生葡萄品种、东亚野生种、欧洲野生种、栽培品种和种间杂交葡萄品种。全文中都使用了这种配色方案。 葡萄系统发育分析SNP的核心集被用于分析葡萄的系统发育和种群结构。用100个非参数bootstraps的最大似然(ML)系统发育分析揭示了北美野生葡萄(WNA,紫色)、东亚野生葡萄(WEA,黄色)、欧洲野生种(WEU,红色)和栽培葡萄(CEU,深绿色,图 2a和补充图 3)的独特单系进化分枝。这种拓扑结构支持之前的报道,即含有北美野生种(新世界物种)的葡萄属进化枝是欧亚葡萄的姊妹种,含有东亚野生种、欧洲野生、和栽培种。 在欧亚进化枝中,欧洲野生种是栽培种的姊妹进化枝,而反过来,作为整体进化枝,它是东亚野生进化枝的姐妹。仔细检查系统发育节点(bootstrap值≥75)后,发现在野生北美分支和东亚野生葡萄分支中分别至少存在两个和五个较小的组。大多数种间杂交的葡萄品种(HYB,浅绿色)被分为两个单独的簇,主要反映了它们在北美野生种、东亚野生种和栽培种之间的各种杂交背景。 ML系统发育树还揭示了一些种质被归类为其他具有不同遗传背景的葡萄属进化枝,这表明在样品收集过程中可能基于形态学对种质进行错误分类。例如,来自加那利群岛圣安德列斯的TA-5901最初被认为是 V. vinifer。系统发育树表明,这种标本很可能是来自北非的 * V. sylvestris*。另一个例子是TA-6147,在加州大学戴维斯分校的收藏中被鉴定为 V. sylvestris (2006年引入KIT)。以前使用质体标记的研究对这种分类法产生了怀疑,因为它与北美野生种聚集在一起。我们的系统树证实了这种错误识别,并显示TA-6147可能是 Vitis acerifolia 的样品。
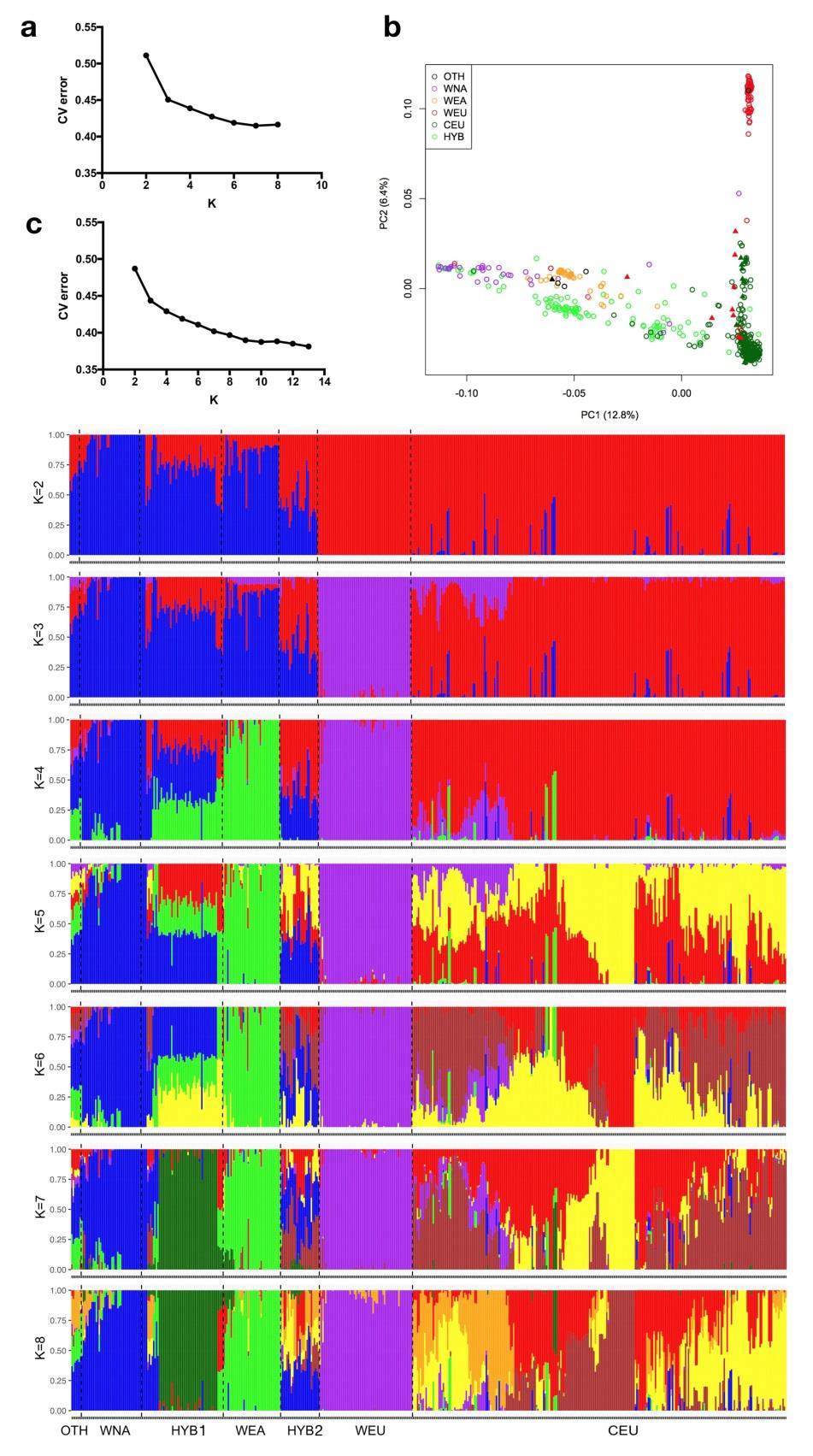
图2 主要葡萄的系统发育和种群结构。a 从具有100个非参数引导程序的全基因组SNP推断出的葡萄种质的最大似然系统树。爬山虎(Parthenocissus tricuspidata,波士顿常春藤)被用作外群。b使用全基因组SNP数据对主要葡萄种的前三个成分进行PCA图。c主要葡萄的种群结构,由ADMIXTURE进行估算。每种颜色代表一个祖先群体。每个种质都由一个条形表示,条形中每个有色段的长度表示该祖先群体贡献的比例。 葡萄种群结构分析主成分分析(PCA)显示了主要葡萄品种之间的大量遗传多样性,前三个主成分分别解释了总遗传变异的13.4%,8.1%和4.7%(图 2b)。 PC1将欧亚种(WEU和CEU)与东亚野生种和北美野生种分离,这表明后两种葡萄品种在遗传背景上比欧亚种具有更多的相似性。基于群体混合的基于模型的分析结果进一步支持了这一发现(图 2c,K=3;补充图 5a,c)。PC2显然将欧洲野生种与驯养的葡萄种分开,而PC3将北美野生种与东亚野生种区分开来(图 2b)。这些主要的葡萄种类之间的区别在种群混合图中也很明显(图 2c,K=5;补充图 5a和5c)。 额外的PCA分析表明,种间杂交的葡萄品种散布在其他葡萄品种之间(补充图 5b)。它们在图中的相对位置反映了每种亲本葡萄属物种的基因组影响水平。在K处的混合图中 = 4(补充图 5c),很明显,大多数种间杂交葡萄簇1(HYB1)受到了北美野生,东亚野生和栽培的葡萄树种的遗传贡献,而种间杂交葡萄簇2 (HYB2)仅从北美野生和栽培的葡萄树种获得遗传贡献。此结果与ML系统发育树相符(补充图 4)。即使大多数栽培的葡萄种在PCA图中都紧密地聚集在一起,但它们显示出明显的高遗传异质性模式,如种群混合分析所示(图 2c,K=6;补充图 5c,K=8)。
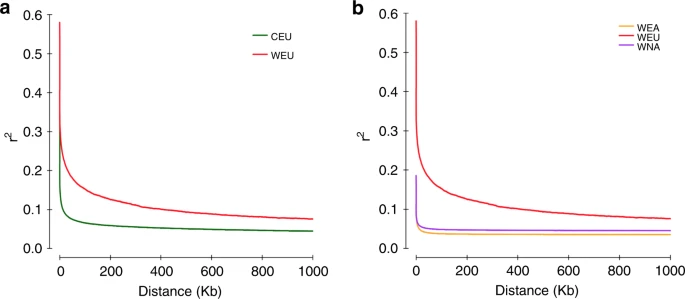
连锁不平衡(LD,表示为r2)模式的表征对于植物的遗传学研究至关重要。之前使用小规模的遗传标记在欧洲野生种和栽培种的研究中,得出的LD衰减并不一致:一些研究显示了10-20 kb的相对小的LD,而其他则找到了28-485kb的大LD区段。利用全基因组SNP,作者发现达到最大平均r2一半的LD中,欧洲野生种为2.9kb,而栽培种则为350bp。这些参数比以前报道的野生大豆(〜27 kb)、野生稻(20 kb)和野生玉米(22 kb)中发现的参数要小得多。 欧洲野生种与栽培种LD的相对较慢的衰减是一致的与以前的研究结果,但要认识随着欧洲葡萄野生种群的多样化,这种差异可能会缩小。与欧洲野生种相比,作者还发现北美野生种和东亚野生种的LD在300 bp内迅速衰减至平均r2为0.1 (图 3b)。
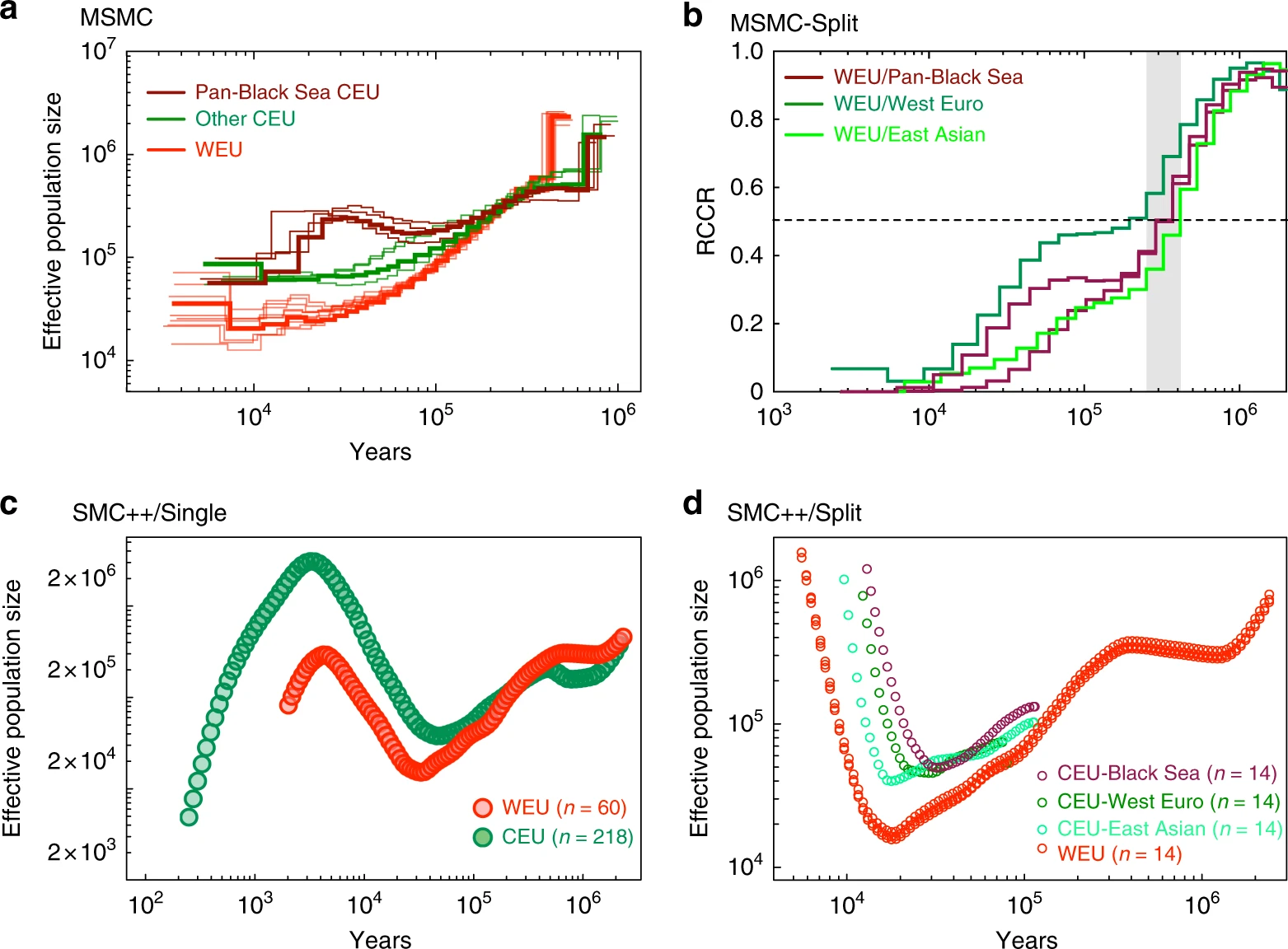
图3 主要葡萄品种的连锁不平衡的衰减。a 欧洲野生葡萄和栽培葡萄品种基因组连锁不平衡的衰退。b 现存野生葡萄品种葡萄基因组连锁不平衡的衰退 WEU和CEU葡萄树的群体历史驯化一年生作物的群体历史(如玉米和非洲水稻),其特征是在驯化期间,估计有效种群规模(Ne)连续收缩和扩张。然而,在具有营养繁殖能力的多年生植物栽培葡萄中没有观察到这种模式(尤其是Ne扩张)。 为了重新探讨这个问题,作者对欧洲野生种和栽培种的 phased SNP data 使用了multiple sequentially Markovian coalescent(MSMC)模型进行分析(图4a)。通过假设产生时间为3年,中性突变率为每年 5.4×10−9,结果被缩放到真实时间。 如图4a所示,欧洲野生种(红线)经历了从 400Kya 处的最高点(Ne≈600,000)到 10 Kya 处的最低点(Ne≈20,000)的持续下降。世界上大多数栽培种(绿线)也观察到了类似的群体统计模式,10–20 Kya 处出现了最低的Ne≈60,000。有趣的是,来自泛黑海地区(褐红色系、高加索地区和黑海西海岸)的栽培葡萄在 30–70 Kya 左右表现出独特的轻微Ne扩张(Ne≈150,000至Ne≈300,000),随后在 10-30Kya 左右出现中度Ne收缩(Ne≈300,000至Ne≈ 60,000)。图4a还显示,在大约 250–300 Kya 的相似时期,所有栽培葡萄的种群开始与欧洲野生葡萄不同。通过分析MSMC split 分析中成对群体的8个单倍型(图4b),作者发现相对交叉合并率达到了0.5,表明在欧洲野生葡萄和栽培葡萄(所有品系)之间分开的时间约为250–400 Kya。
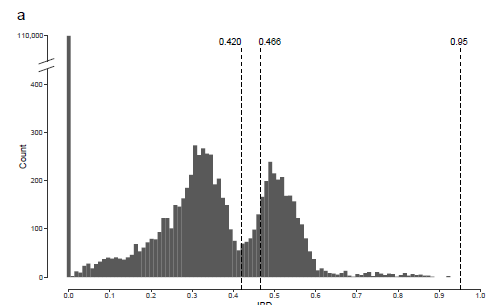
图4 欧洲野生葡萄和栽培葡萄的群体历史。a MSMC分析的从104年到106年前的WEU和CEU葡萄群体统计历史。每一行代表来自两个单独材料的四个单倍型。每个组都突出了一个例子。对于CEU葡萄,来自泛黑海地区的葡萄品种与其他葡萄品种相比产生了不同的群体统计学历史。b 基于相对交叉聚结率(RCCR)的WEU和CEU葡萄各个亚组的MSMC拆分分析。c WEU(n=60)和CEU(n=218)的群体统计学历史从102到106年前来自SMC ++的单种群分析。d WEU(n=14)和CEU的各种亚组(n=14)的葡萄从104到106年前,根据SMC++分裂分析 由于研究中MSMC的功能在10 Kya之前的预测中受到很大限制,因此作者使用SMC++ 探索了具有无阶段SNP数据的葡萄的近期群体统计历史。综合考虑MSMC和SMC++的结果,作者认为泛黑海品种自从欧洲野生种开始就经历了独特的群体历史。此外,自上次冰河时代以来,所有葡萄都可能经历了至少一个瓶颈。由于在葡萄研究中很难鉴定出真正的祖先群体,因此作者建议对所有涉及欧洲野生葡萄的分析都应谨慎解释。 系谱分析由于营养繁殖和有性繁殖的并存,很难从基因组数据准确地重建葡萄谱系。尽管如此,作者分析了472个葡萄品种之间的IBD关系模式。成对的 Vitis 比较得出的IBD值的直方图是双峰(bimodal)的。成对IBD值均不超过0.95(定义克隆性的经验界限)。通过将IBD的临界值定义为0.420(分隔两种模式的最低值),我们发现335 Vitis一级亲缘关系(71.0%)与至少一个其他种质相关。当对43对已确认的葡萄亲子对采用最低成对IBD值(0.466)时,292份葡萄种质(61.9%)与至少一份其他种质保持一级关系。
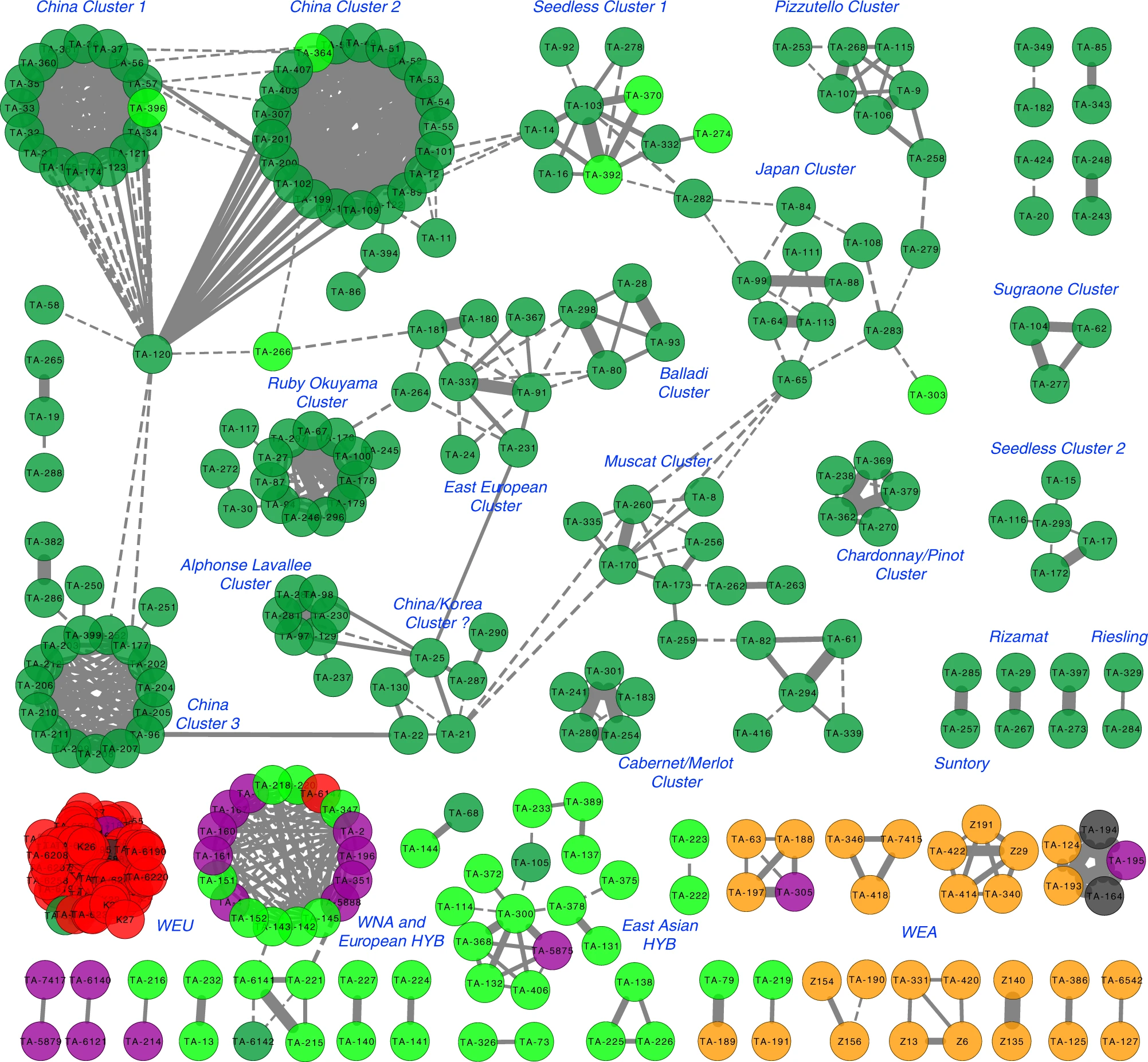
附图6a,IBD的双峰分布 作者使用Cytoscape进行了谱系网络可视化。图中显示,大多数一级关系都在相同的主要类别的种质之间。欧洲野生葡萄藤形成一个紧凑的独立集群。相比之下,北美野生种与欧洲种间杂交种高度相关,这与在19世纪欧洲葡萄园中广泛使用北美野生葡萄来对抗霉菌和害虫的历史相吻合。东亚野生种形成了一个松散连接的簇,这意味着尚未充分探索东亚野生种来创造杂交品种。驯化的葡萄品种形成了16个离散的簇。中国品种形成了三个高度连接的紧凑型集群,这些集群通过关键节点品种“Jingxiu”链接在一起(图 5)。相比之下,欧洲品种形成了松散连接的分散集群。例如,独立的集群显示了“Cabernet Sauvignon/Merlot”和“Chardonnay/Pinot”的一级关系,对应于先前的报道。另一个例子是形成两个单独的无核葡萄簇,一个以“Thompson Seedless”为中心,另一个以“Emperor”为中心。驯化的葡萄品种网络表明,数千年前开始的欧洲驯化的葡萄品种的繁殖历史非常复杂。
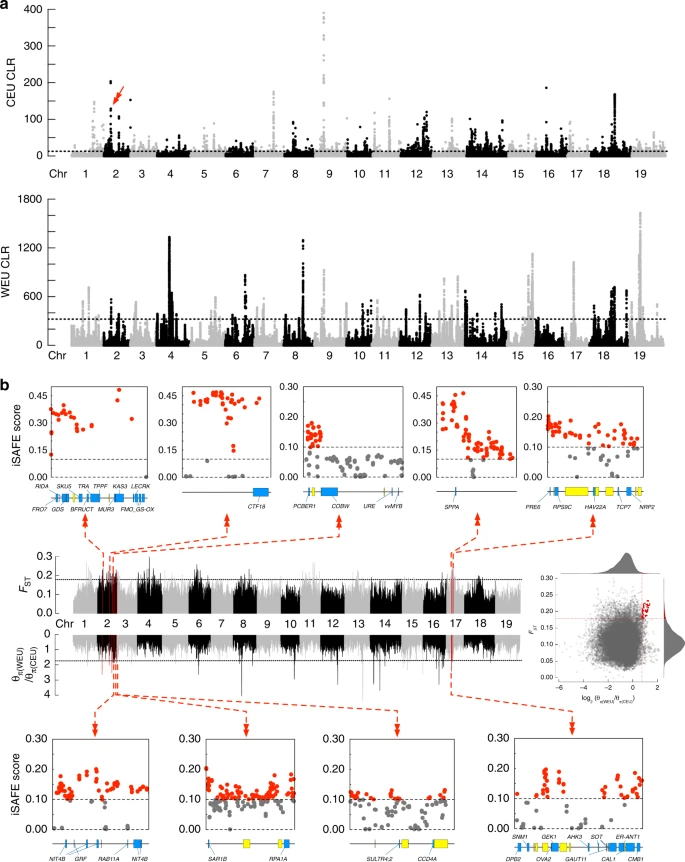
图5 WEU和CEU中的选择信号在葡萄基因组中,全局核苷酸多样性(π)与全局SNP和indel密度密切相关(图1b)。随后的分析表明,欧洲野生葡萄的多态性程度(π=3.50×10−3;S.D.=1.84×10−3)低于驯化品种(π=5.49×10−3;S.D.=1.91×10−3)。欧洲野生葡萄品种与驯化葡萄品种的遗传多样性差异在一些独立的葡萄研究中得到了很好的注意。考虑到野生欧洲葡萄的Ne显著减少(图4),其多样性水平的降低可能是由于在最近的几年里,分散的本地栖息地和增加的近亲繁殖的结果。 尽管有这个缺点,我们还是通过鉴定在CLR分析中得分最高为0.5%的区域(长度约1 kb)调查了欧洲野生和驯化的葡萄树基因组中的潜在选择性信号(图 6a)。驯化的葡萄树基因组中的2119个选择性清除区域包含1016个候选基因,而野生葡萄树基因组中的2120个选择性清除区域包含348个候选基因。鉴定的基因数量比以前的研究更大(1016对308,共有28个公共基因; 348对88,无共有基因)。特别是,先前确定的第2号染色体上的花性决定基因座(4.91–5.05 Mb 或 4.88–5.04 Mb)被发现在驯化的葡萄树基因组中包含15个重要的CLR信号。在欧洲野生的和驯化的葡萄树的基因组中没有发现重叠的选择性清除区域,并且在两个列表中都发现了18个基因。1016个驯化的葡萄候选基因的GO富集在免疫反应、细胞死亡调节、果糖代谢过程、质膜和囊泡运输的GO类别中显示出重要的功能性表示。有趣的是,发现茉莉酸酯诱导的细胞死亡与葡萄中的非生物防御反应有关。此外,花青素的囊泡运输起着浆果皮肤着色重要作用和浆果葡萄藤催熟。该结果表明,驯化的葡萄树可能主要是由于果糖含量更高,成熟时间各异以及种植面积更广而选择的。相比之下,GO富集的348个野生欧洲葡萄候选基因在大分子代谢,芳香族化合物生物合成和茉莉酸代谢途径的GO类别中显示出重要的功能代表,表明对生物和非生物胁迫具有抗性的自然选择。
图6 葡萄基因组中的选择性扫描区域。a 由SweeD计算得出的欧洲野生和驯养葡萄树中整个基因组的CLR分数。虚线标记了顶部0.5%的区域。红色箭头指示假定的性别确定区域。b从FST和π推断驯化期间的选择性扫描区域统计数据和每个区域的SNP热点受正在进行的选择扫描的影响。每个选择性扫描区域由基因组中的红色垂直线表示。每个选择性扫描区域的放大视图由一条水平线表示,在其上绘制了注释的(蓝色方框)和未知的(黄色方框)基因。iSAFE图中的每个点代表该区域内36个SNP(步长为18个SNP)的平均iSAFE得分。虚线显示了经验意义的临界值。在第一个iSAFE图中,该区域与假定的性别确定区域重叠 |
 、
、【本文地址】
| 今日新闻 |
| 推荐新闻 |
| 专题文章 |